

Actin-dependent activation of serum response factor in T cells by the viral oncoprotein tip
- PMID: 22385615
- PMCID: PMC3310822
- DOI: 10.1186/1478-811X-10-5
Actin-dependent activation of serum response factor in T cells by the viral oncoprotein tip
Abstract
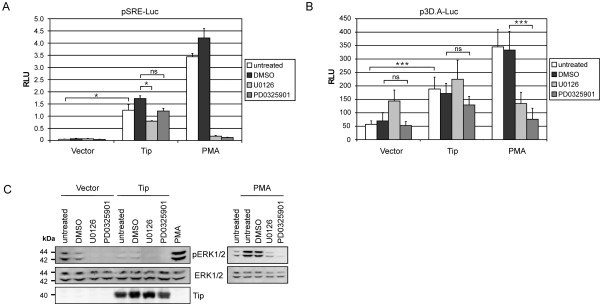
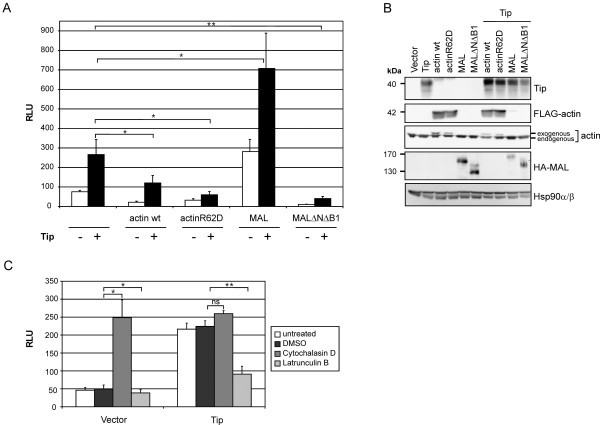
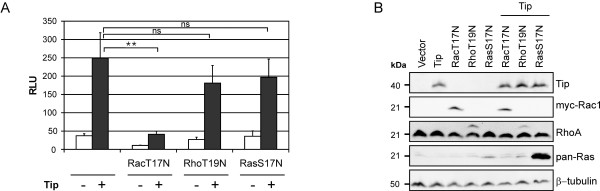
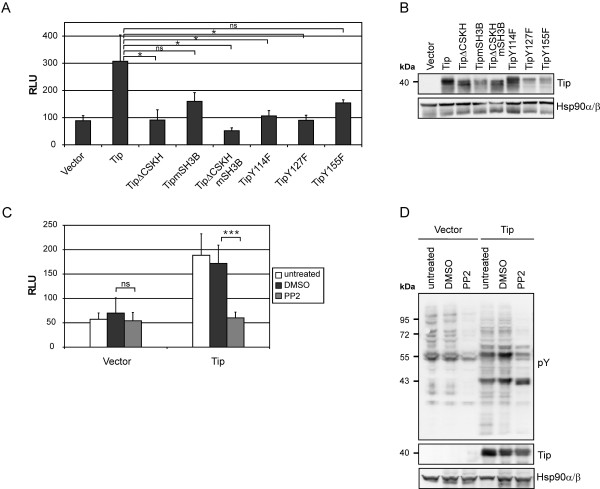
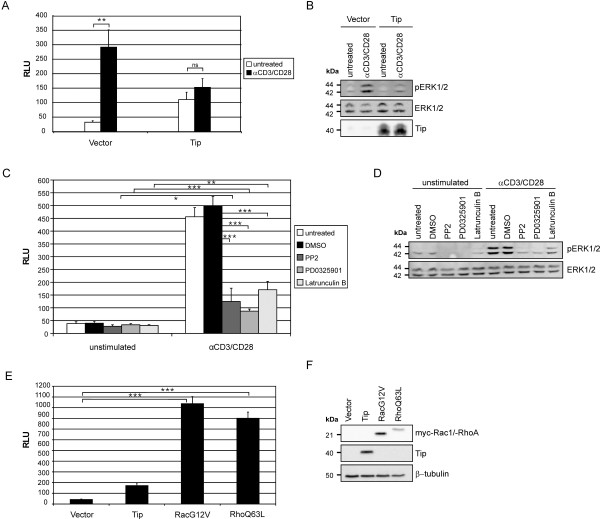
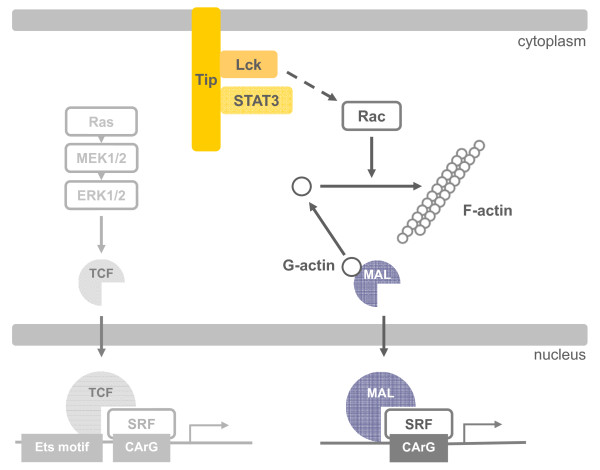
Serum response factor (SRF) acts as a multifunctional transcription factor regulated by mutually exclusive interactions with ternary complex factors (TCFs) or myocardin-related transcription factors (MRTFs). Binding of Rho- and actin-regulated MRTF:SRF complexes to target gene promoters requires an SRF-binding site only, whereas MAPK-regulated TCF:SRF complexes in addition rely on flanking sequences present in the serum response element (SRE). Here, we report on the activation of an SRE luciferase reporter by Tip, the viral oncoprotein essentially contributing to human T-cell transformation by Herpesvirus saimiri. SRE activation in Tip-expressing Jurkat T cells could not be attributed to triggering of the MAPK pathway. Therefore, we further analyzed the contribution of MRTF complexes. Indeed, Tip also activated a reporter construct responsive to MRTF:SRF. Activation of this reporter was abrogated by overexpression of a dominant negative mutant of the MRTF-family member MAL. Moreover, enrichment of monomeric actin suppressed the Tip-induced reporter activity. Further upstream, the Rho-family GTPase Rac, was found to be required for MRTF:SRF reporter activation by Tip. Initiation of this pathway was strictly dependent on Tip's ability to interact with Lck and on the activity of this Src-family kinase. Independent of Tip, T-cell stimulation orchestrates Src-family kinase, MAPK and actin pathways to induce SRF. These findings establish actin-regulated transcription in human T cells and suggest its role in viral oncogenesis.
Figures
Similar articles
-
Megakaryoblastic leukemia 1, a potent transcriptional coactivator for serum response factor (SRF), is required for serum induction of SRF target genes.Mol Cell Biol. 2003 Sep;23(18):6597-608. doi: 10.1128/MCB.23.18.6597-6608.2003. Mol Cell Biol. 2003. PMID: 12944485 Free PMC article.
-
Rho-actin signaling to the MRTF coactivators dominates the immediate transcriptional response to serum in fibroblasts.Genes Dev. 2014 May 1;28(9):943-58. doi: 10.1101/gad.239327.114. Epub 2014 Apr 14. Genes Dev. 2014. PMID: 24732378 Free PMC article.
-
Growth hormone regulates ternary complex factors and serum response factor associated with the c-fos serum response element.J Biol Chem. 1997 Oct 10;272(41):25951-8. doi: 10.1074/jbc.272.41.25951. J Biol Chem. 1997. PMID: 9325329
-
MKLs: co-factors of serum response factor (SRF) in neuronal responses.Int J Biochem Cell Biol. 2012 Sep;44(9):1444-7. doi: 10.1016/j.biocel.2012.05.008. Epub 2012 May 22. Int J Biochem Cell Biol. 2012. PMID: 22626970 Review.
-
SRF'ing and SAP'ing - the role of MRTF proteins in cell migration.J Cell Sci. 2018 Oct 11;131(19):jcs218222. doi: 10.1242/jcs.218222. J Cell Sci. 2018. PMID: 30309957 Free PMC article. Review.
Cited by
-
EZH2 regulates neuroepithelium structure and neuroblast proliferation by repressing p21.Open Biol. 2016 Apr;6(4):150227. doi: 10.1098/rsob.150227. Epub 2016 Apr 20. Open Biol. 2016. PMID: 27248655 Free PMC article.
-
Network Modeling Approach to Predict Myofibroblast Differentiation.Cell Mol Bioeng. 2014 Sep;7(3):446-459. doi: 10.1007/s12195-014-0344-9. Epub 2014 Jul 6. Cell Mol Bioeng. 2014. PMID: 33072223 Free PMC article.
-
Difference in F-actin depolymerization induced by toxin B from the Clostridium difficile strain VPI 10463 and toxin B from the variant Clostridium difficile serotype F strain 1470.Toxins (Basel). 2013 Jan 11;5(1):106-19. doi: 10.3390/toxins5010106. Toxins (Basel). 2013. PMID: 23344455 Free PMC article.
-
Dasatinib inhibits TGFβ-induced myofibroblast differentiation through Src-SRF Pathway.Eur J Pharmacol. 2015 Dec 15;769:134-42. doi: 10.1016/j.ejphar.2015.11.008. Epub 2015 Nov 6. Eur J Pharmacol. 2015. PMID: 26548624 Free PMC article.
-
Effect of mycalolides isolated from a marine sponge Mycale aff. nullarosette on actin in living cells.Sci Rep. 2019 May 17;9(1):7540. doi: 10.1038/s41598-019-44036-2. Sci Rep. 2019. PMID: 31101864 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
