

Single-molecule analysis of the microtubule cross-linking protein MAP65-1 reveals a molecular mechanism for contact-angle-dependent microtubule bundling
- PMID: 22385851
- PMCID: PMC3283812
- DOI: 10.1016/j.bpj.2012.01.008
Single-molecule analysis of the microtubule cross-linking protein MAP65-1 reveals a molecular mechanism for contact-angle-dependent microtubule bundling
Abstract
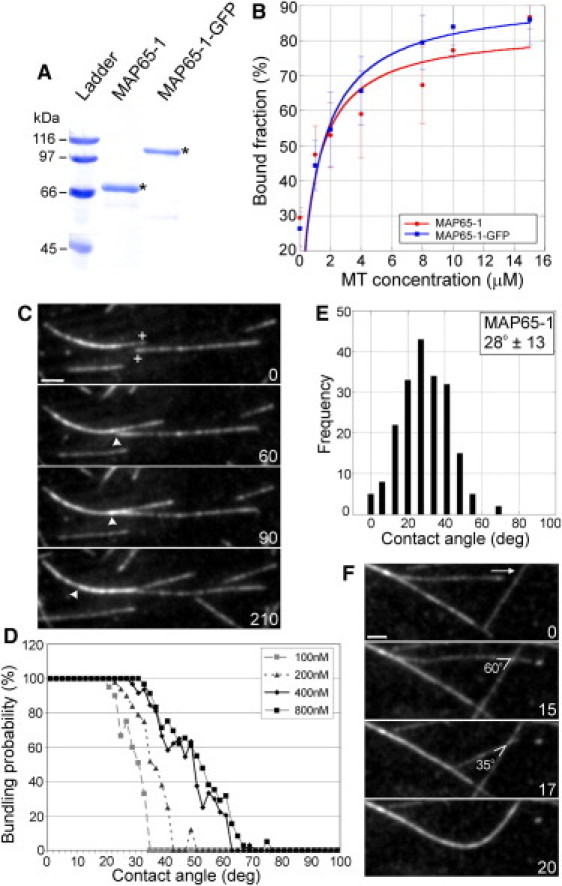
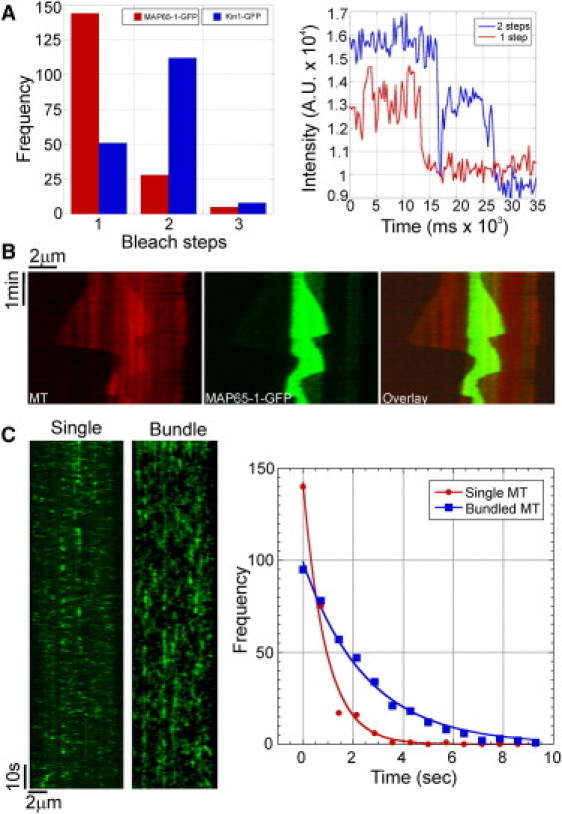
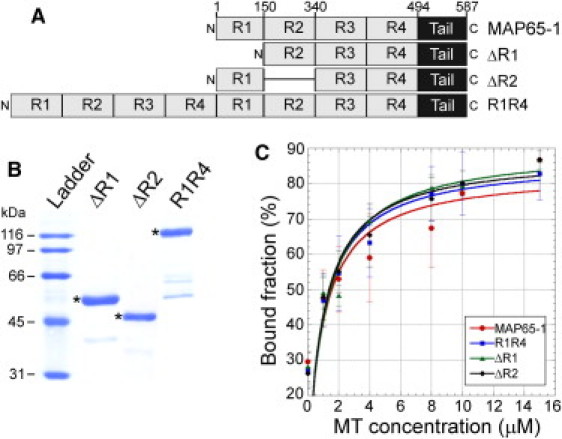
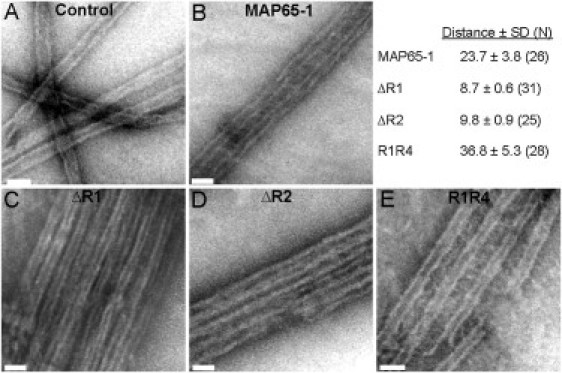
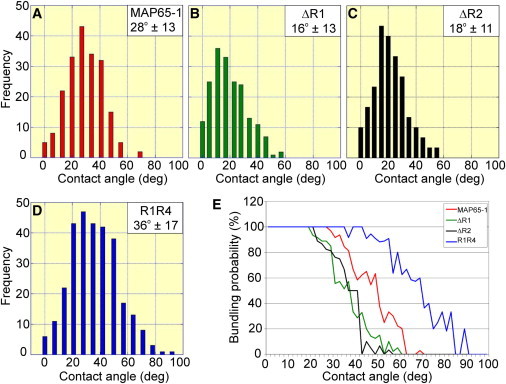
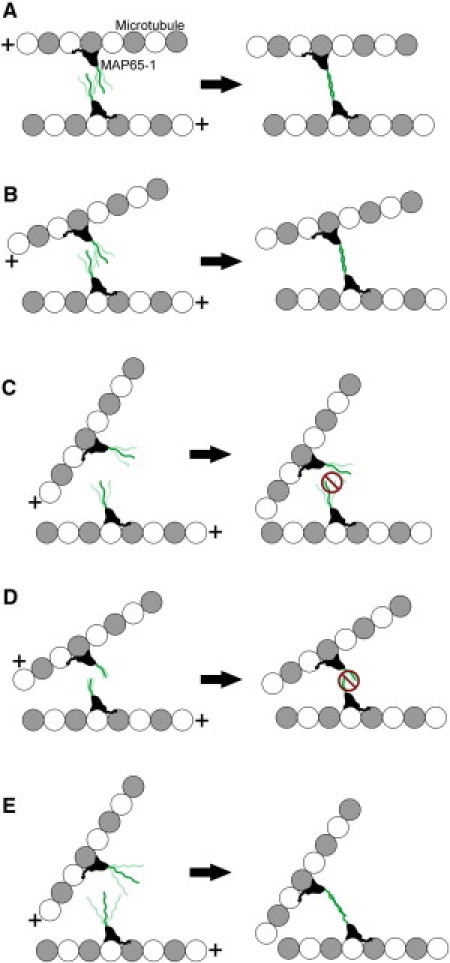
Bundling of microtubules (MTs) is critical for the formation of complex MT arrays. In land plants, the interphase cortical MTs form bundles specifically following shallow-angle encounters between them. To investigate how cells select particular MT contact angles for bundling, we used an in vitro reconstitution approach consisting of dynamic MTs and the MT-cross-linking protein MAP65-1. We found that MAP65-1 binds to MTs as monomers and inherently targets antiparallel MTs for bundling. Dwell-time analysis showed that the affinity of MAP65-1 for antiparallel overlapping MTs is about three times higher than its affinity for single MTs and parallel overlapping MTs. We also found that purified MAP65-1 exclusively selects shallow-angle MT encounters for bundling, indicating that this activity is an intrinsic property of MAP65-1. Reconstitution experiments with mutant MAP65-1 proteins with different numbers of spectrin repeats within the N-terminal rod domain showed that the length of the rod domain is a major determinant of the range of MT bundling angles. The length of the rod domain also determined the distance between MTs within a bundle. Together, our data show that the rod domain of MAP65-1 acts both as a spacer and as a structural element that specifies the MT encounter angles that are conducive for bundling.
Copyright © 2012 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Wasteneys G.O., Ambrose J.C. Spatial organization of plant cortical microtubules: close encounters of the 2D kind. Trends Cell Biol. 2009;19:62–71. - PubMed
-
- Shaw S.L., Kamyar R., Ehrhardt D.W. Sustained microtubule treadmilling in Arabidopsis cortical arrays. Science. 2003;300:1715–1718. - PubMed
-
- Nakamura M., Ehrhardt D.W., Hashimoto T. Microtubule and katanin-dependent dynamics of microtubule nucleation complexes in the acentrosomal Arabidopsis cortical array. Nat. Cell Biol. 2010;12:1064–1070. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
