

Protein synthesis during sleep consolidates cortical plasticity in vivo
- PMID: 22386312
- PMCID: PMC3350782
- DOI: 10.1016/j.cub.2012.02.016
Protein synthesis during sleep consolidates cortical plasticity in vivo
Abstract
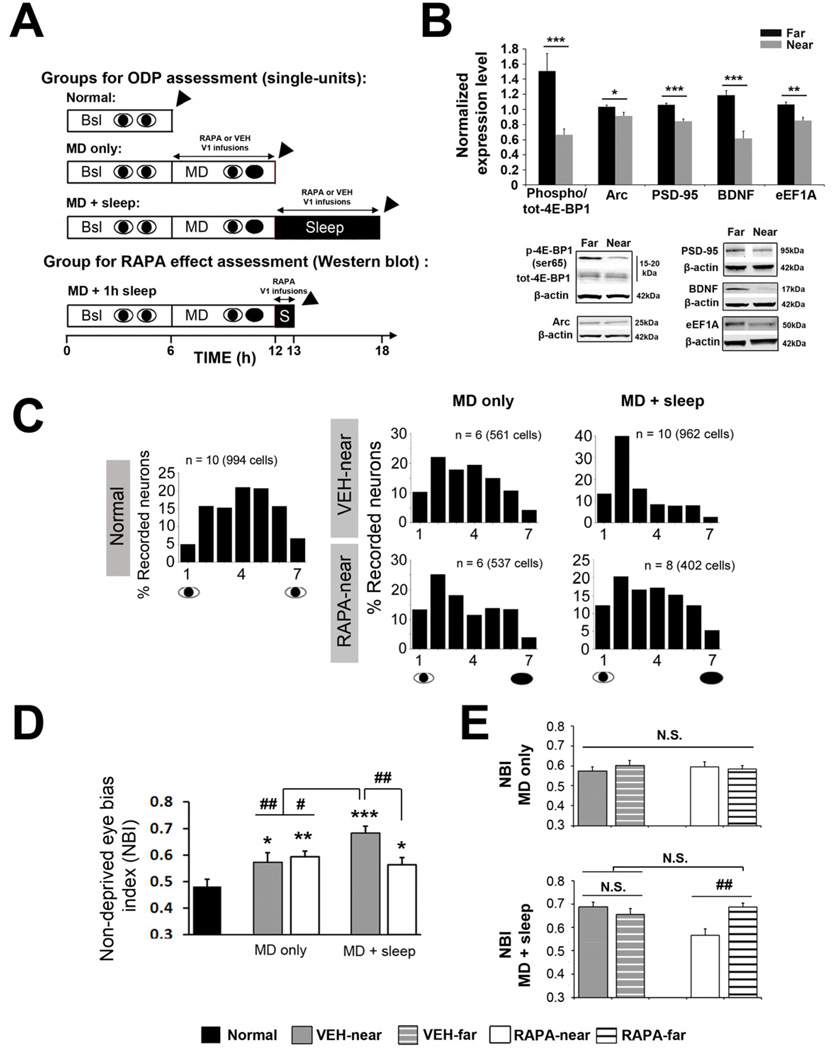
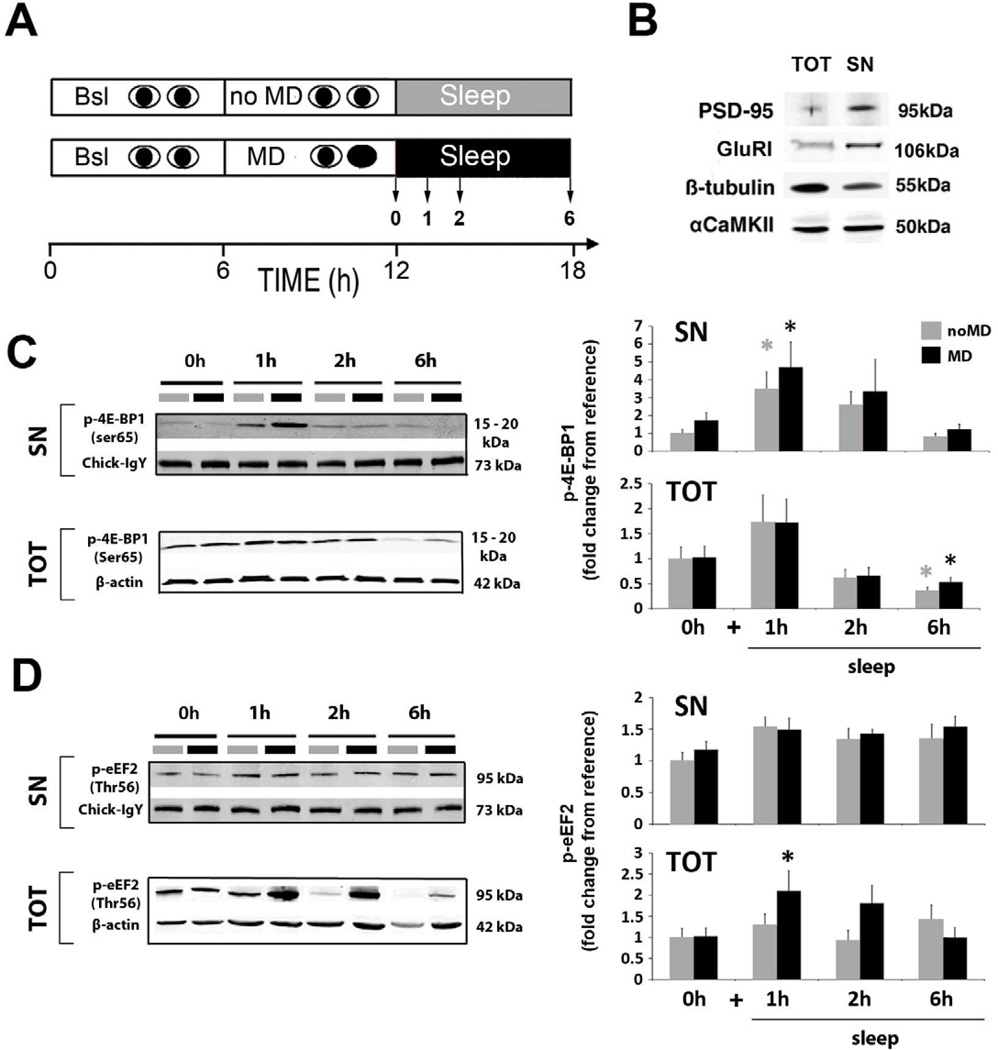
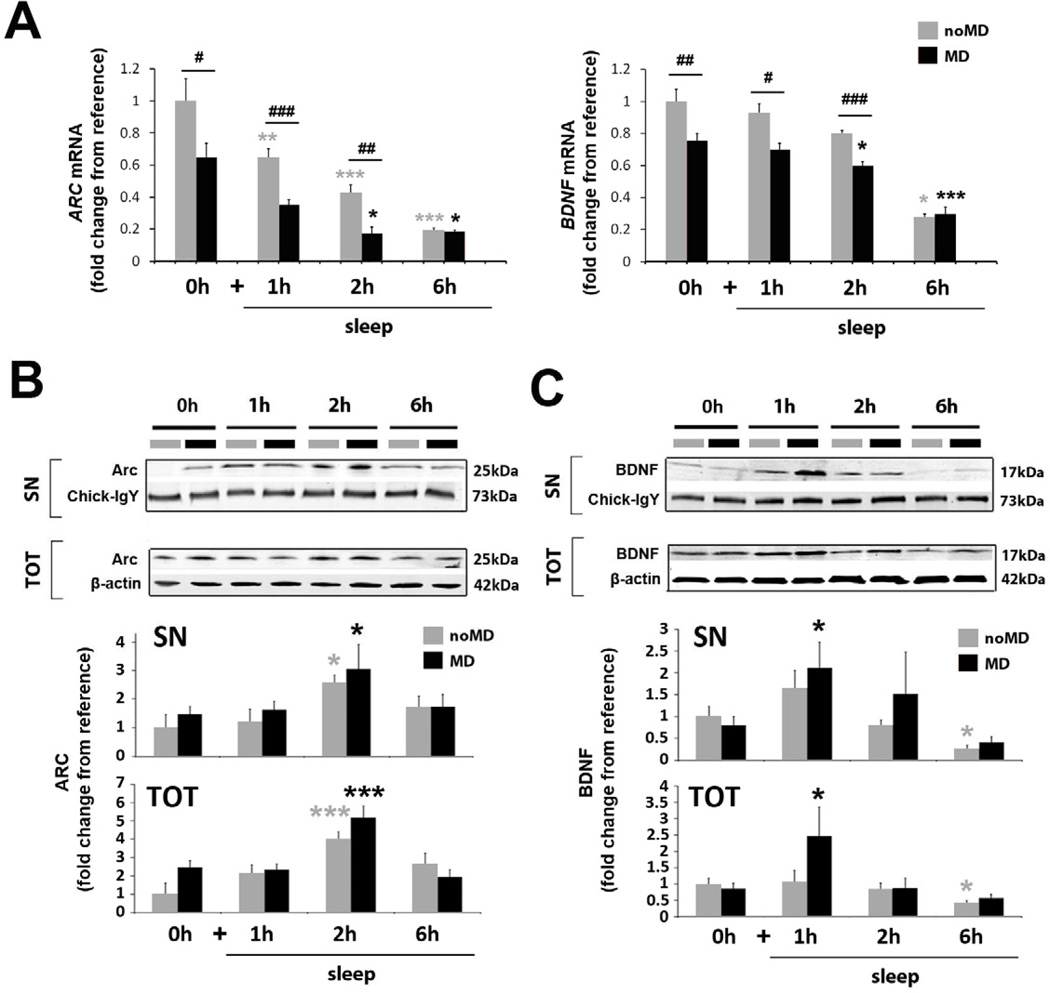
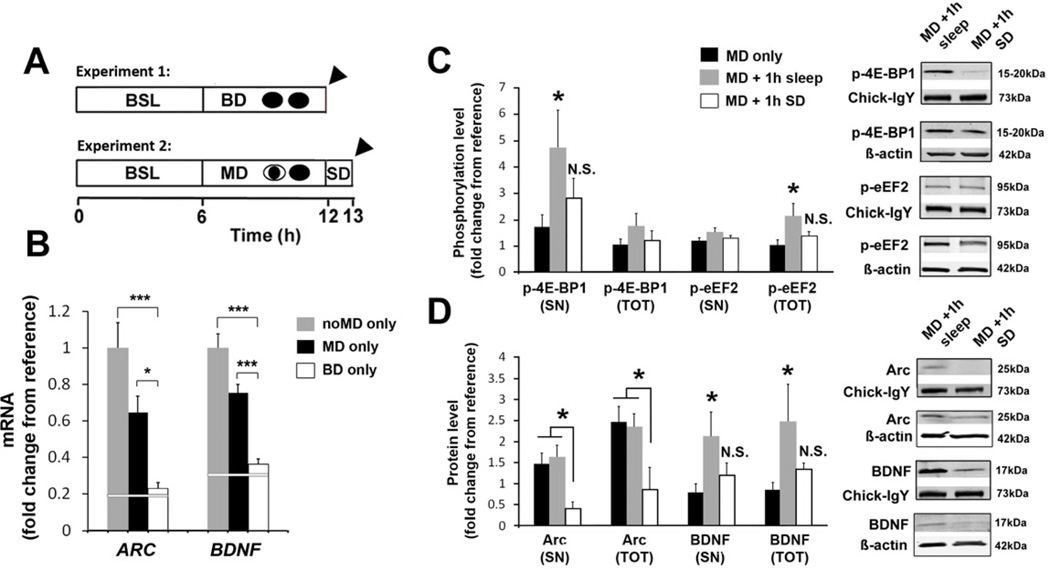
Sleep consolidates experience-dependent brain plasticity, but the precise cellular mechanisms mediating this process are unknown [1]. De novo cortical protein synthesis is one possible mechanism. In support of this hypothesis, sleep is associated with increased brain protein synthesis [2, 3] and transcription of messenger RNAs (mRNAs) involved in protein synthesis regulation [4, 5]. Protein synthesis in turn is critical for memory consolidation and persistent forms of plasticity in vitro and in vivo [6, 7]. However, it is unknown whether cortical protein synthesis in sleep serves similar functions. We investigated the role of protein synthesis in the sleep-dependent consolidation of a classic form of cortical plasticity in vivo (ocular dominance plasticity, ODP; [8, 9]) in the cat visual cortex. We show that intracortical inhibition of mammalian target of rapamycin (mTOR)-dependent protein synthesis during sleep abolishes consolidation but has no effect on plasticity induced during wakefulness. Sleep also promotes phosphorylation of protein synthesis regulators (i.e., 4E-BP1 and eEF2) and the translation (but not transcription) of key plasticity related mRNAs (ARC and BDNF). These findings show that sleep promotes cortical mRNA translation. Interruption of this process has functional consequences, because it abolishes the consolidation of experience in the cortex.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
References
-
- Frank MG, Benington JH. The role of sleep in memory consolidation and brain plasticity: dream or reality? Neuroscientist. 2006;12:477–488. - PubMed
-
- Ramm P, Smith CT. Rates of cerebral protein synthesis are linked to slow wave sleep in the rat. Physiol. Behav. 1990;48:749–753. - PubMed
-
- Nakanishi H, Sun Y, Nakamura RK, Mori K, Ito M, Suda S, Namba H, Storch FI, Dang TP, Mendelson TW, et al. Positive correlations between cerebral protein synthesis rates and deep sleep in Macaca mulatta. Eur. J. Neurosci. 1997;9:271–279. - PubMed
-
- Cirelli C, Gutierrez CM, Tononi G. Extensive and divergent effects of sleep and wakefulness on brain gene expression. Neuron. 2004;41:35–43. - PubMed
-
- Mackiewicz M, Shockley KR, Romer MA, Galante RJ, Zimmerman JE, Naidoo N, Baldwin DA, Jensen ST, Churchill GA, Pack AI. Macromolecule biosynthesis: a key function of sleep. Physiol. Genomics. 2007;31:441–457. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- F32 EY017766/EY/NEI NIH HHS/United States
- F31 NS067935/NS/NINDS NIH HHS/United States
- T32 HL007953/HL/NHLBI NIH HHS/United States
- F32-EY017766/EY/NEI NIH HHS/United States
- R01-EY 019002/EY/NEI NIH HHS/United States
- T32 GM007517/GM/NIGMS NIH HHS/United States
- R00 EY021503/EY/NEI NIH HHS/United States
- R01 EY019022/EY/NEI NIH HHS/United States
- F31-NS067935/NS/NINDS NIH HHS/United States
- T32 EY007143/EY/NEI NIH HHS/United States
- K99 EY021503/EY/NEI NIH HHS/United States
- F32 EY006831/EY/NEI NIH HHS/United States
- T32-GM07517/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Miscellaneous
