

Coding region polyadenylation generates a truncated tRNA synthetase that counters translation repression
- PMID: 22386318
- PMCID: PMC3615456
- DOI: 10.1016/j.cell.2012.02.018
Coding region polyadenylation generates a truncated tRNA synthetase that counters translation repression
Abstract
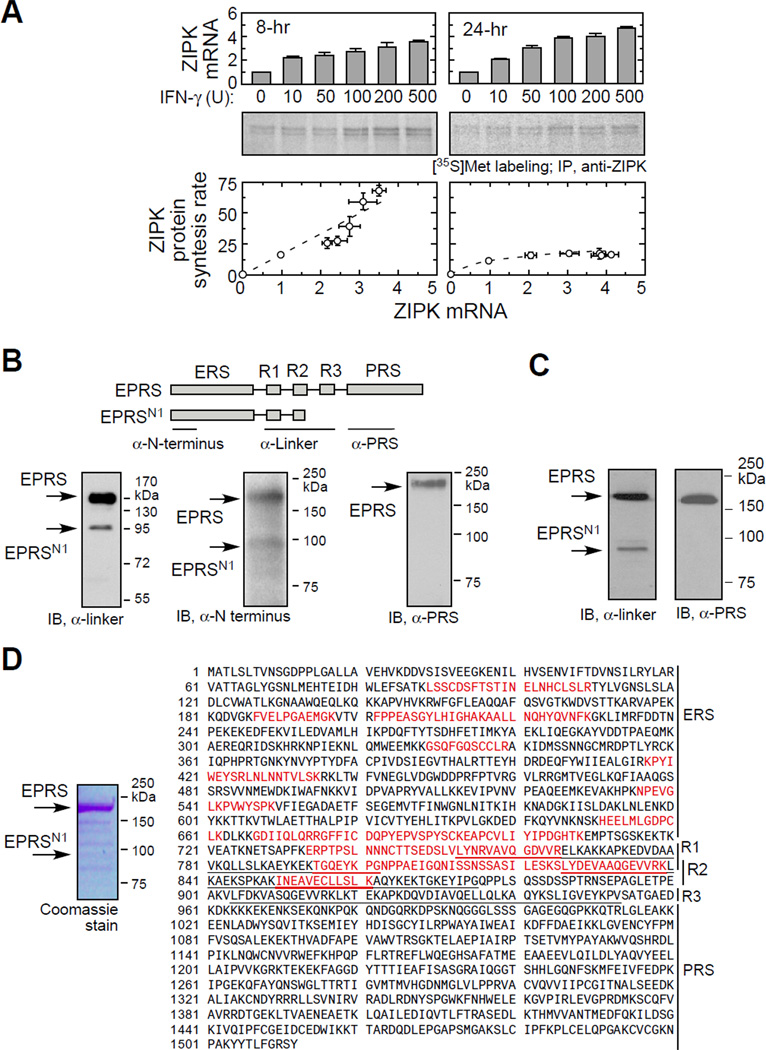
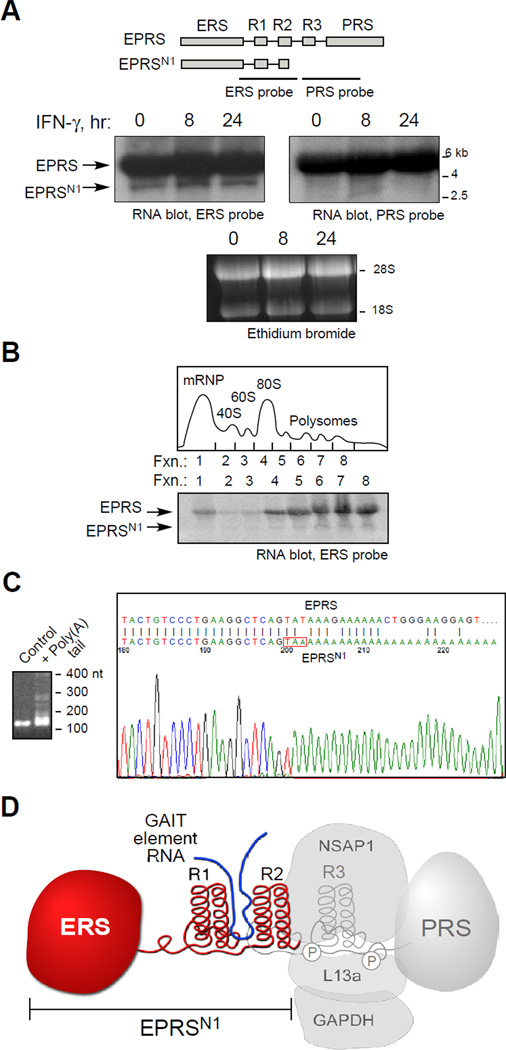
Posttranscriptional regulatory mechanisms superimpose "fine-tuning" control upon "on-off" switches characteristic of gene transcription. We have exploited computational modeling with experimental validation to resolve an anomalous relationship between mRNA expression and protein synthesis. The GAIT (gamma-interferon-activated inhibitor of translation) complex repressed VEGF-A synthesis to a low, constant rate independent of VEGF-A mRNA expression levels. Dynamic model simulations predicted an inhibitory GAIT-element-interacting factor to account for this relationship and led to the identification of a truncated form of glutamyl-prolyl tRNA synthetase (EPRS), a GAIT constituent that mediates binding to target transcripts. The truncated protein, EPRS(N1), shields GAIT-element-bearing transcripts from the inhibitory GAIT complex, thereby dictating a "translational trickle" of GAIT target proteins. EPRS(N1) mRNA is generated by polyadenylation-directed conversion of a Tyr codon in the EPRS-coding sequence to a stop codon (PAY(∗)). Genome-wide analysis revealed multiple candidate PAY(∗) targets, including the authenticated target RRM1, suggesting a general mechanism for production of C terminus-truncated regulatory proteins.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures





Comment in
-
A molecular doorstop ensures a trickle through translational repression.Cell. 2012 Mar 30;149(1):13-5. doi: 10.1016/j.cell.2012.03.010. Cell. 2012. PMID: 22464317
References
-
- Ahmed YF, Gilmartin GM, Hanly SM, Nevins JR, Greene WC. The HTLV-I Rex response element mediates a novel form of mRNA polyadenylation. Cell. 1991;64:727–737. - PubMed
-
- Anderson S, Bankier AT, Barrell BG, de Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F, et al. Sequence and organization of the human mitochondrial genome. Nature. 1981;290:457–465. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
