

CpG methylation recruits sequence specific transcription factors essential for tissue specific gene expression
- PMID: 22387149
- PMCID: PMC3371161
- DOI: 10.1016/j.bbagrm.2012.02.014
CpG methylation recruits sequence specific transcription factors essential for tissue specific gene expression
Abstract
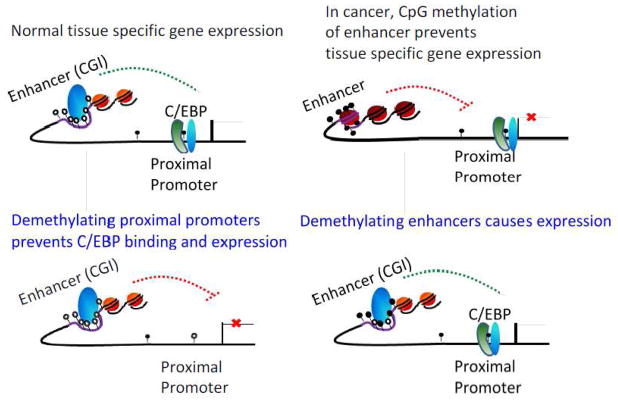
CG methylation is an epigenetically inherited chemical modification of DNA found in plants and animals. In mammals it is essential for accurate regulation of gene expression and normal development. Mammalian genomes are depleted for the CG dinucleotide, a result of the chemical deamination of methyl-cytosine in CG resulting in TpG. Most CG dinucleotides are methylated, but ~15% are unmethylated. Five percent of CGs cluster into ~20,000 regions termed CG islands (CGI) which are generally unmethylated. About half of CGIs are associated with housekeeping genes. In contrast, the gene body, repeats and transposable elements in which CGs are generally methylated. Unraveling the epigenetic machinery operating in normal cells is important for understanding the epigenetic aberrations that are involved in human diseases including cancer. With the advent of high-throughput sequencing technologies, it is possible to identify the CG methylation status of all 30million unique CGs in the human genome, and monitor differences in distinct cell types during differentiation and development. Here we summarize the present understanding of DNA methylation in normal cells and discuss recent observations that CG methylation can have an effect on tissue specific gene expression. We also discuss how aberrant CG methylation can lead to cancer. This article is part of a Special Issue entitled: Chromatin in time and space.
Published by Elsevier B.V.
Figures



References
-
- Bird A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002;16:6–21. - PubMed
-
- Lister R, Pelizzola M, Dowen RH, Hawkins RD, Hon G, Tonti-Filippini J, Nery JR, Lee L, Ye Z, Ngo QM, Edsall L, Antosiewicz-Bourget J, Stewart R, Ruotti V, Millar AH, Thomson JA, Ren B, Ecker JR. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature. 2009;462:315–322. - PMC - PubMed
-
- Illingworth RS, Bird AP. CpG islands--'a rough guide'. FEBS Lett. 2009;583:1713–1720. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
