

A sensor-adaptor mechanism for enterovirus uncoating from structures of EV71
- PMID: 22388738
- PMCID: PMC3378640
- DOI: 10.1038/nsmb.2255
A sensor-adaptor mechanism for enterovirus uncoating from structures of EV71
Abstract
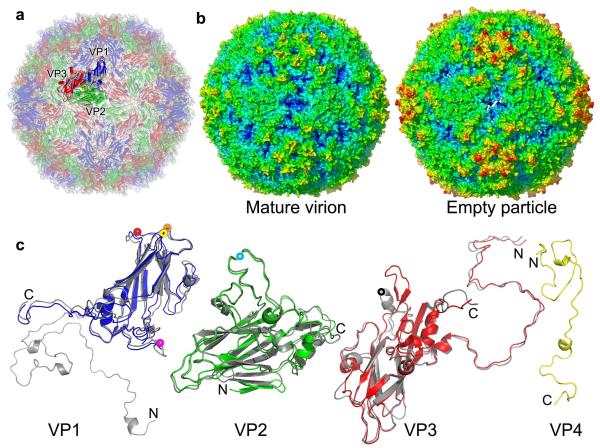
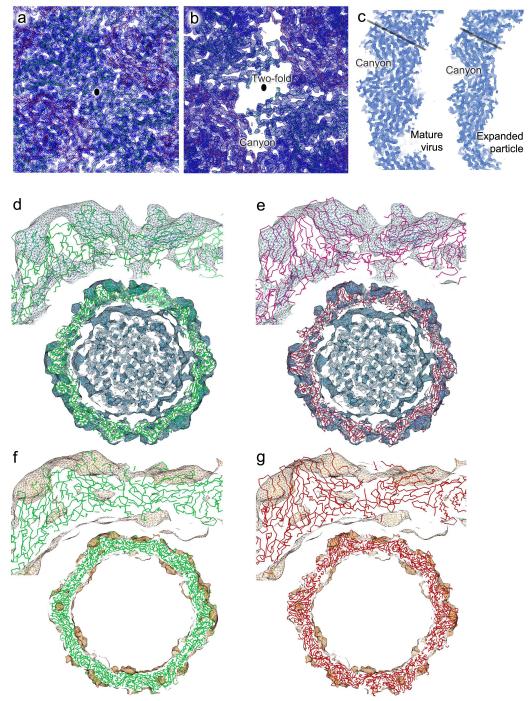
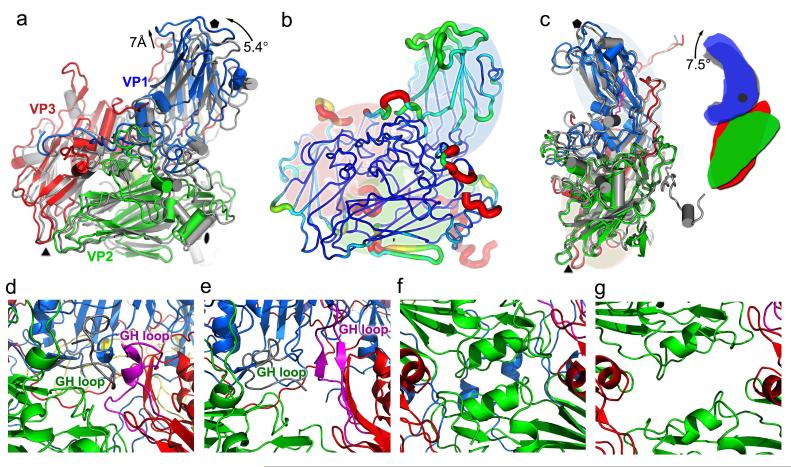
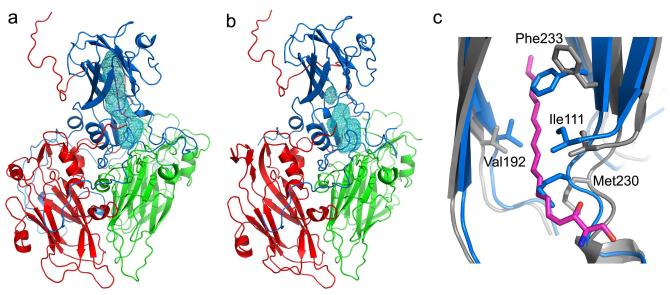
Enterovirus 71 (EV71) is a major agent of hand, foot and mouth disease in children that can cause severe central nervous system disease and death. No vaccine or antiviral therapy is available. High-resolution structural analysis of the mature virus and natural empty particles shows that the mature virus is structurally similar to other enteroviruses. In contrast, the empty particles are markedly expanded and resemble elusive enterovirus-uncoating intermediates not previously characterized in atomic detail. Hydrophobic pockets in the EV71 capsid are collapsed in this expanded particle, providing a detailed explanation of the mechanism for receptor-binding triggered virus uncoating. These structures provide a model for enterovirus uncoating in which the VP1 GH loop acts as an adaptor-sensor for cellular receptor attachment, converting heterologous inputs to a generic uncoating mechanism, highlighting new opportunities for therapeutic intervention.
Figures
Comment in
-
A 3D framework for understanding enterovirus 71.Nat Struct Mol Biol. 2012 Apr 4;19(4):367-8. doi: 10.1038/nsmb.2276. Nat Struct Mol Biol. 2012. PMID: 22472617
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
