

Homeostatic control of recombination is implemented progressively in mouse meiosis
- PMID: 22388890
- PMCID: PMC3319518
- DOI: 10.1038/ncb2451
Homeostatic control of recombination is implemented progressively in mouse meiosis
Abstract
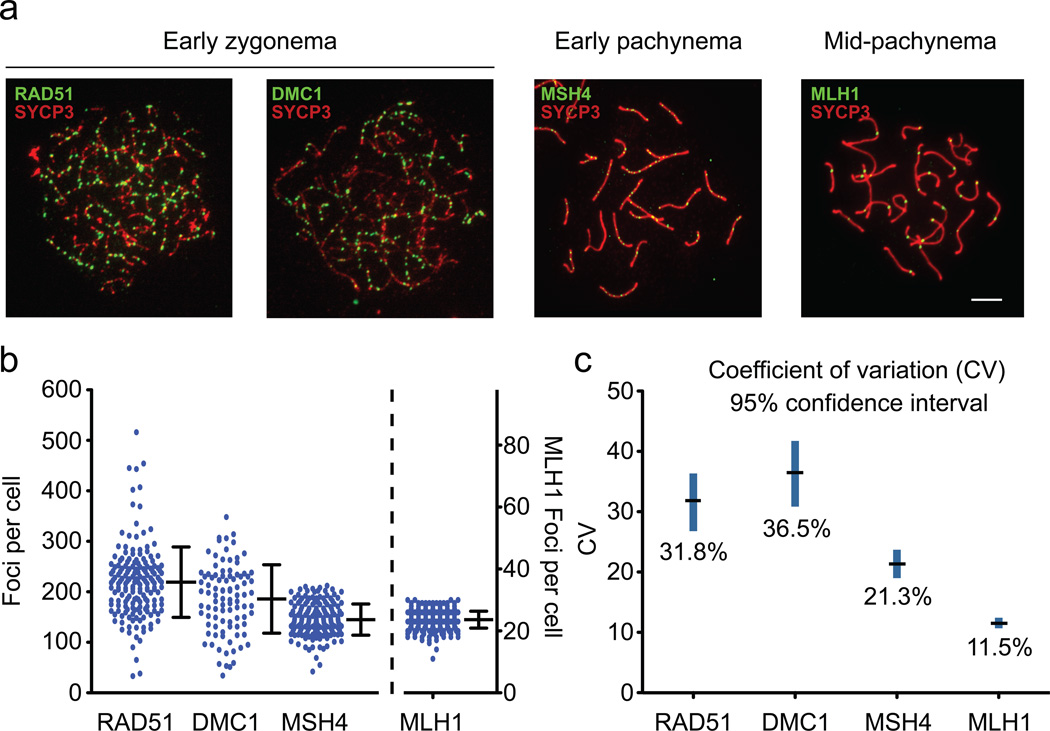
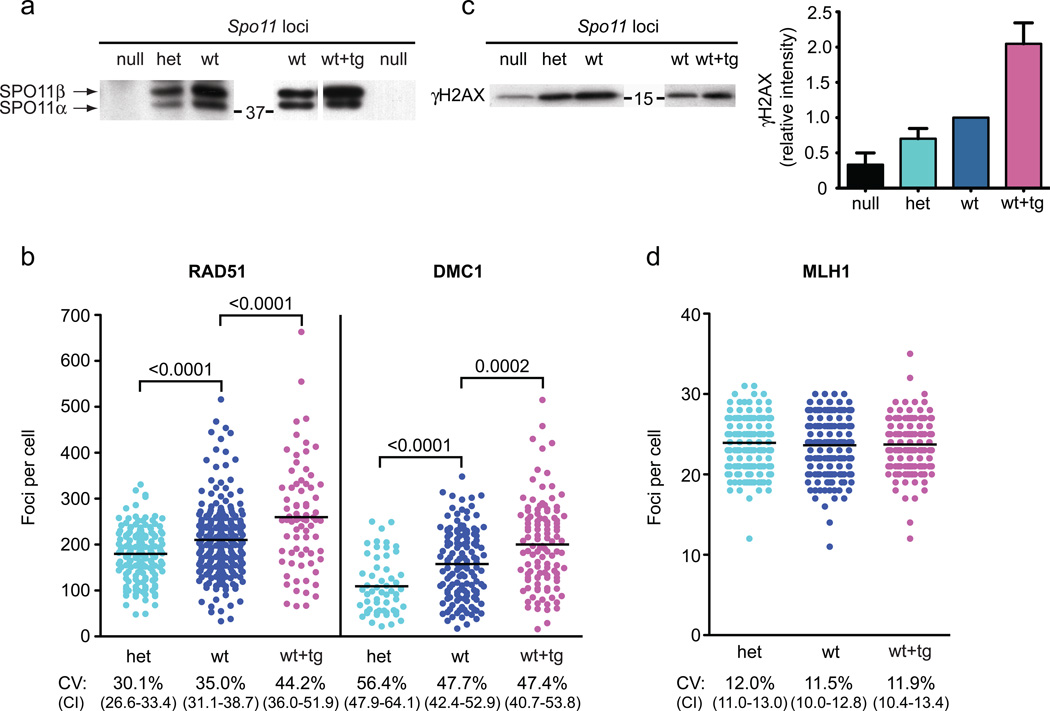
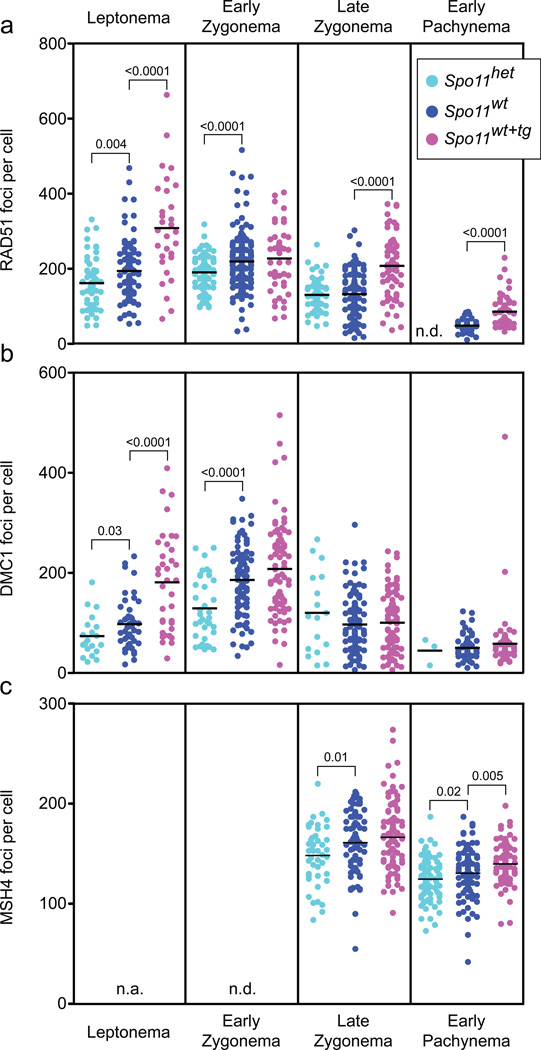
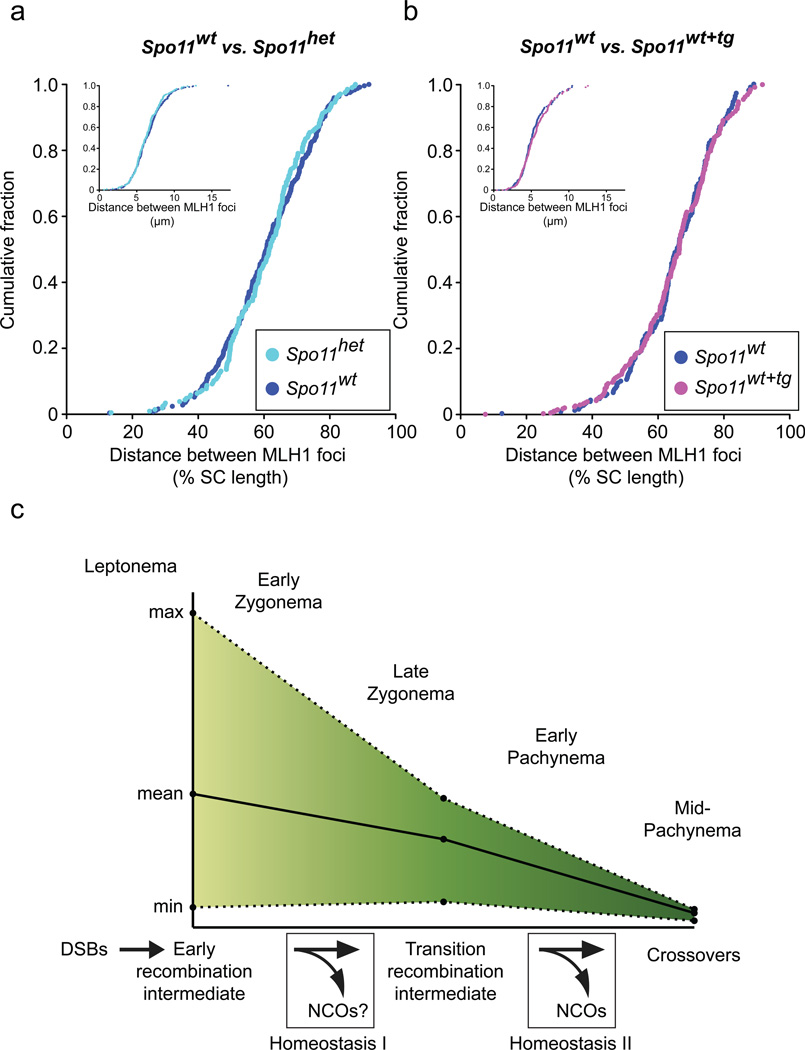
Humans suffer from high rates of fetal aneuploidy, often arising from the absence of meiotic crossover recombination between homologous chromosomes. Meiotic recombination is initiated by double-strand breaks (DSBs) generated by the SPO11 transesterase. In yeast and worms, at least one buffering mechanism, crossover homeostasis, maintains crossover numbers despite variation in DSB numbers. We show here that mammals exhibit progressive homeostatic control of recombination. In wild-type mouse spermatocytes, focus numbers for early recombination proteins (RAD51, DMC1) were highly variable from cell to cell, whereas foci of the crossover marker MLH1 showed little variability. Furthermore, mice with greater or fewer copies of the Spo11 gene--with correspondingly greater or fewer numbers of early recombination foci--exhibited relatively invariant crossover numbers. Homeostatic control is enforced during at least two stages, after the formation of early recombination intermediates and later while these intermediates mature towards crossovers. Thus, variability within the mammalian meiotic program is robustly managed by homeostatic mechanisms to control crossover formation, probably to suppress aneuploidy. Meiotic recombination exemplifies how order can be progressively implemented in a self-organizing system despite natural cell-to-cell disparities in the underlying biochemical processes.
Figures
Comment in
-
Robustness in crossover regulation during meiosis.Nat Cell Biol. 2012 Apr 2;14(4):335-7. doi: 10.1038/ncb2474. Nat Cell Biol. 2012. PMID: 22469828
Similar articles
-
FIGNL1-FIRRM is essential for meiotic recombination and prevents DNA damage-independent RAD51 and DMC1 loading.Nat Commun. 2024 Aug 15;15(1):7015. doi: 10.1038/s41467-024-51458-8. Nat Commun. 2024. PMID: 39147779 Free PMC article.
-
Variation in genome-wide levels of meiotic recombination is established at the onset of prophase in mammalian males.PLoS Genet. 2014 Jan 30;10(1):e1004125. doi: 10.1371/journal.pgen.1004125. eCollection 2014 Jan. PLoS Genet. 2014. PMID: 24497841 Free PMC article.
-
Meiotic crossover control by concerted action of Rad51-Dmc1 in homolog template bias and robust homeostatic regulation.PLoS Genet. 2013;9(12):e1003978. doi: 10.1371/journal.pgen.1003978. Epub 2013 Dec 19. PLoS Genet. 2013. PMID: 24367271 Free PMC article.
-
Roles of RecA homologues Rad51 and Dmc1 during meiotic recombination.Cytogenet Genome Res. 2004;107(3-4):201-7. doi: 10.1159/000080598. Cytogenet Genome Res. 2004. PMID: 15467365 Review.
-
Double-stranded DNA breaks and gene functions in recombination and meiosis.Cell Res. 2006 May;16(5):402-12. doi: 10.1038/sj.cr.7310052. Cell Res. 2006. PMID: 16699536 Review.
Cited by
-
DNA methylation restrains transposons from adopting a chromatin signature permissive for meiotic recombination.Genes Dev. 2015 Jun 15;29(12):1256-70. doi: 10.1101/gad.257840.114. Genes Dev. 2015. PMID: 26109049 Free PMC article.
-
Control of Meiotic Crossovers: From Double-Strand Break Formation to Designation.Annu Rev Genet. 2016 Nov 23;50:175-210. doi: 10.1146/annurev-genet-120215-035111. Epub 2016 Sep 14. Annu Rev Genet. 2016. PMID: 27648641 Free PMC article. Review.
-
Meiotic recombination in mammals: localization and regulation.Nat Rev Genet. 2013 Nov;14(11):794-806. doi: 10.1038/nrg3573. Nat Rev Genet. 2013. PMID: 24136506 Review.
-
DNA Helicase Mph1FANCM Ensures Meiotic Recombination between Parental Chromosomes by Dissociating Precocious Displacement Loops.Dev Cell. 2020 May 18;53(4):458-472.e5. doi: 10.1016/j.devcel.2020.04.010. Epub 2020 May 7. Dev Cell. 2020. PMID: 32386601 Free PMC article.
-
Mutation and recombination parameters in zebra finch are similar to those in mammals.bioRxiv [Preprint]. 2025 Feb 17:2024.09.05.611523. doi: 10.1101/2024.09.05.611523. bioRxiv. 2025. Update in: PLoS Genet. 2025 Apr 15;21(4):e1011661. doi: 10.1371/journal.pgen.1011661. PMID: 39282267 Free PMC article. Updated. Preprint.
References
-
- Hassold T, Hall H, Hunt P. The origin of human aneuploidy: where we have been, where we are going. Hum Mol Genet. 2007;16(Spec No. 2):R203–R208. - PubMed
-
- Keeney S. Spo11 and the formation of DNA double-strand breaks in meiosis. In: Egel R, Lankenau D-H, editors. Recombination and Meiosis . Berlin Heidelberg: Springer-Verlag; 2007. pp. 81–123.
-
- Hillers KJ, Villeneuve AM. Chromosome-wide control of meiotic crossing over in C. elegans. Curr Biol. 2003;13:1641–1647. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
