

Native-like aggregates of factor VIII are immunogenic in von Willebrand factor deficient and hemophilia a mice
- PMID: 22388918
- PMCID: PMC3774159
- DOI: 10.1002/jps.23091
Native-like aggregates of factor VIII are immunogenic in von Willebrand factor deficient and hemophilia a mice
Abstract
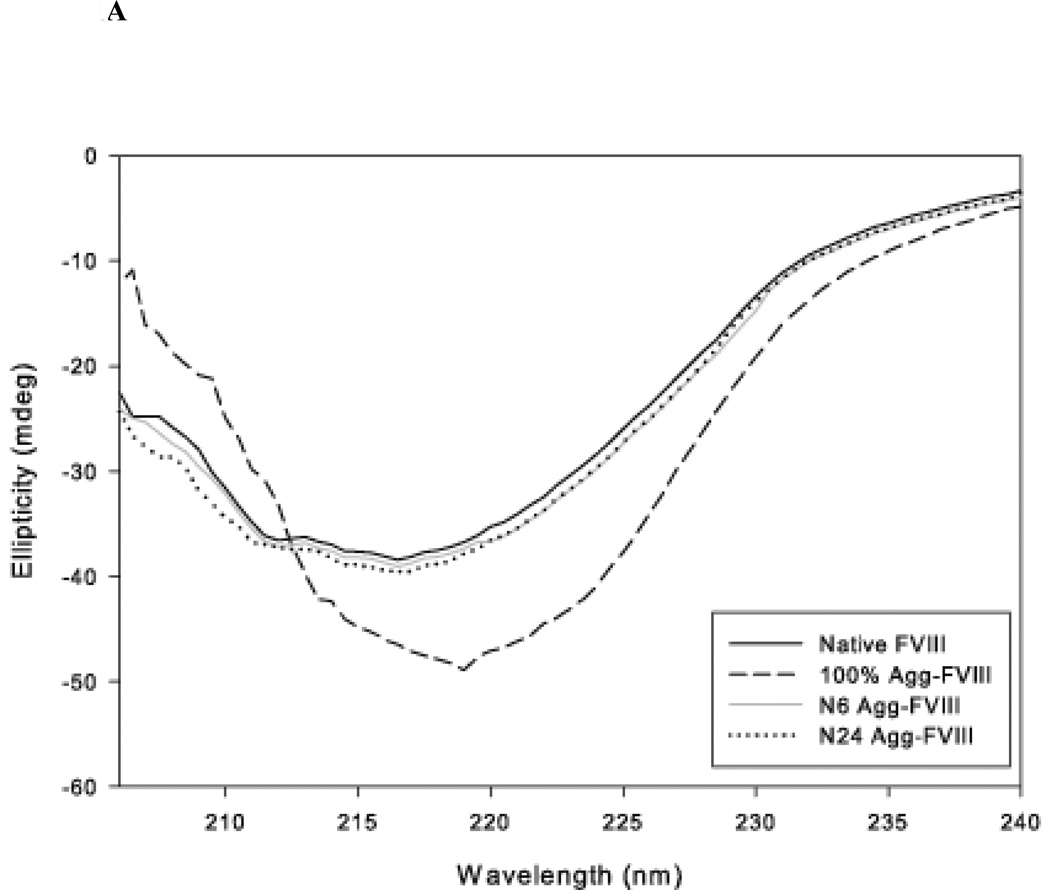
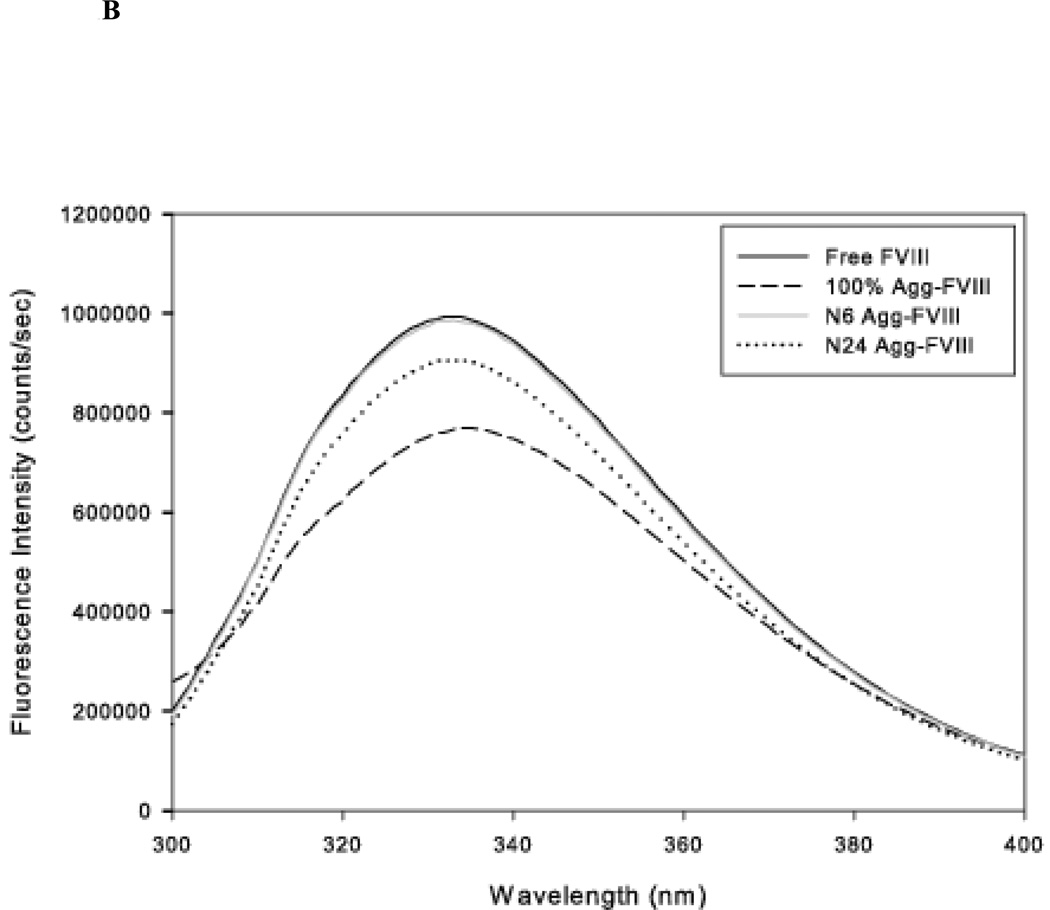
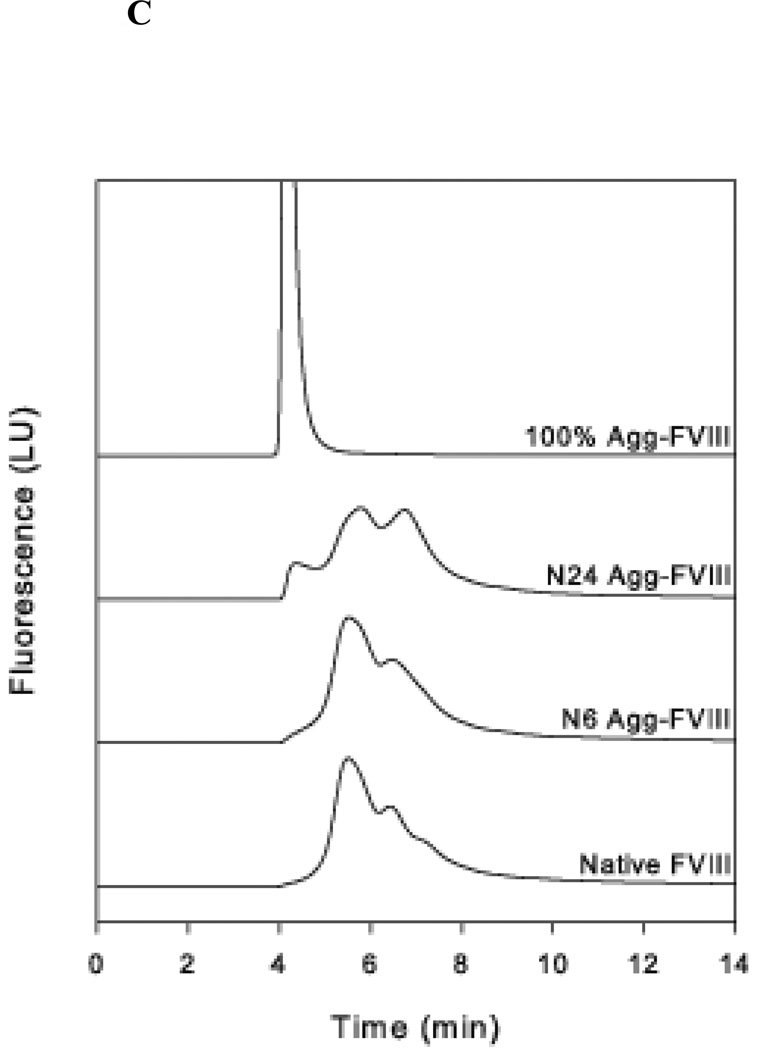
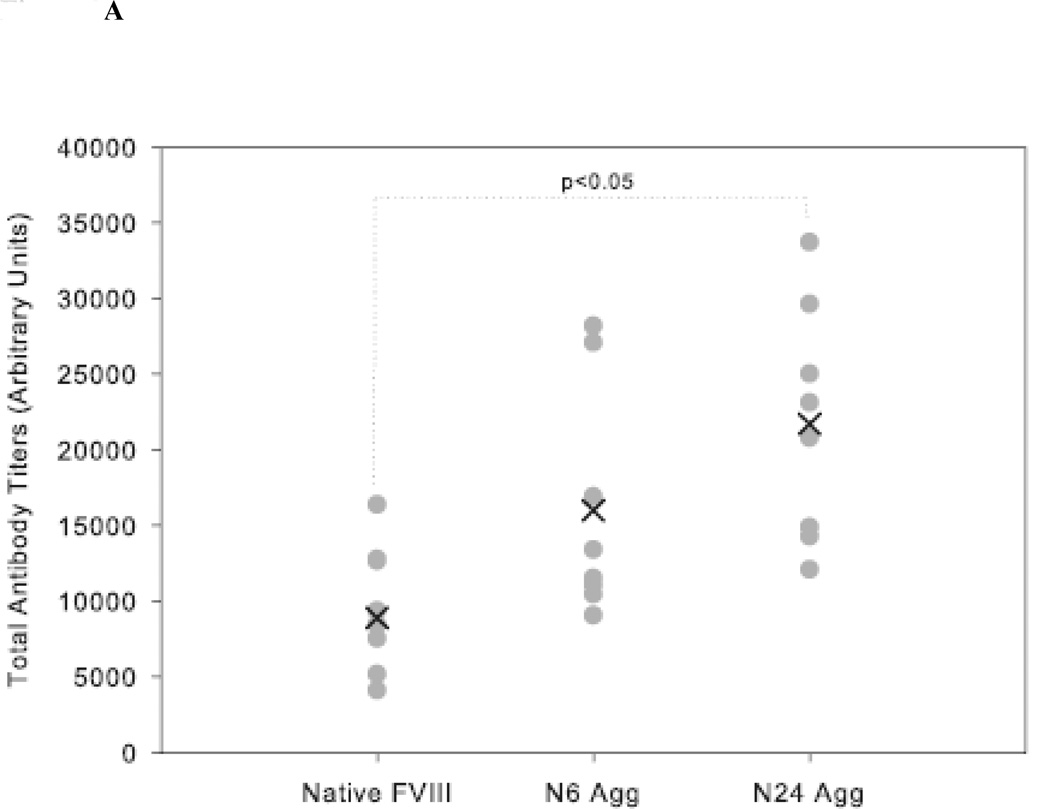
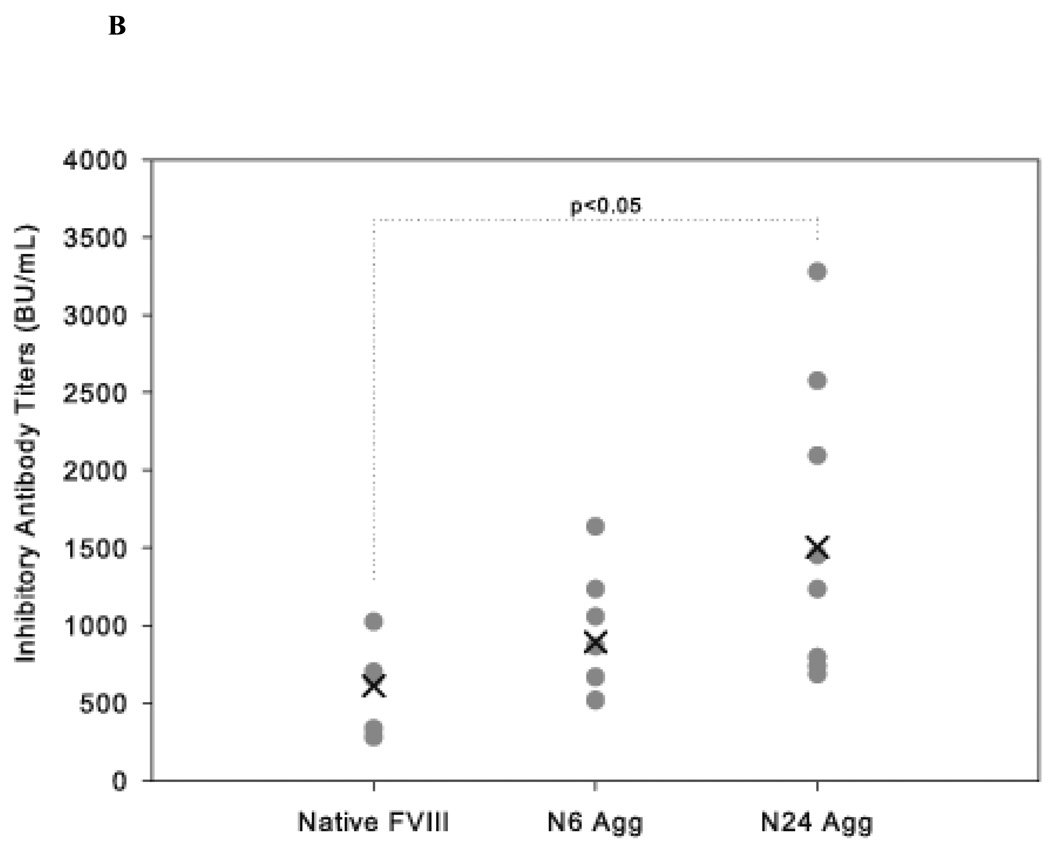
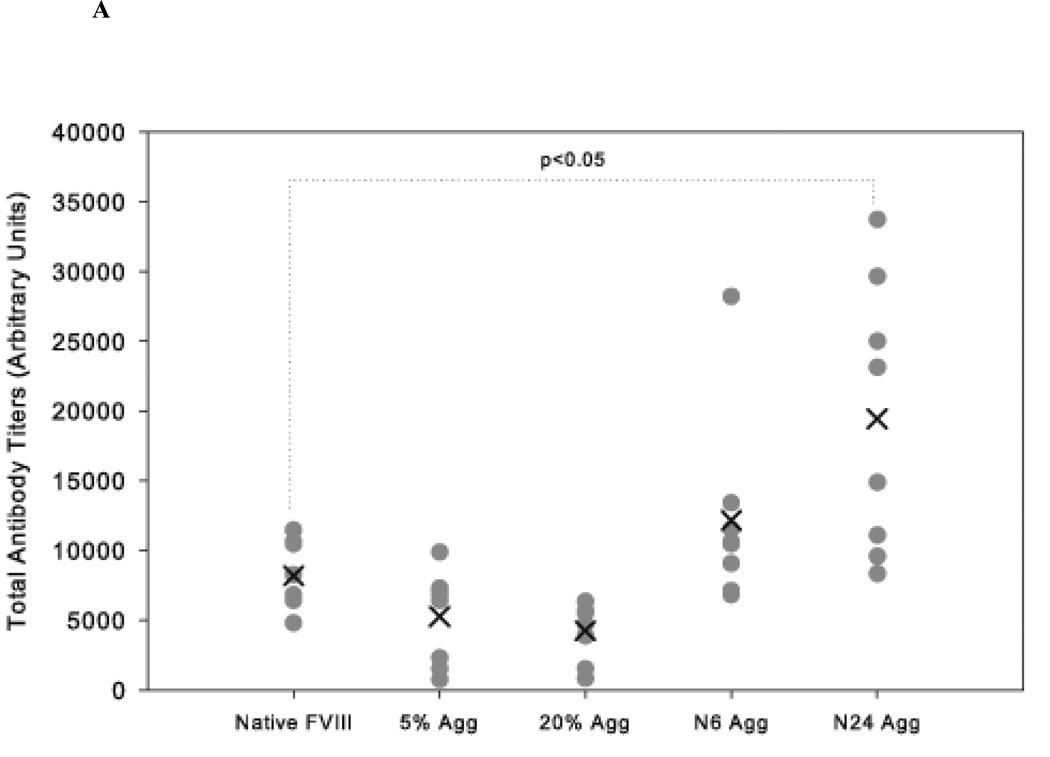
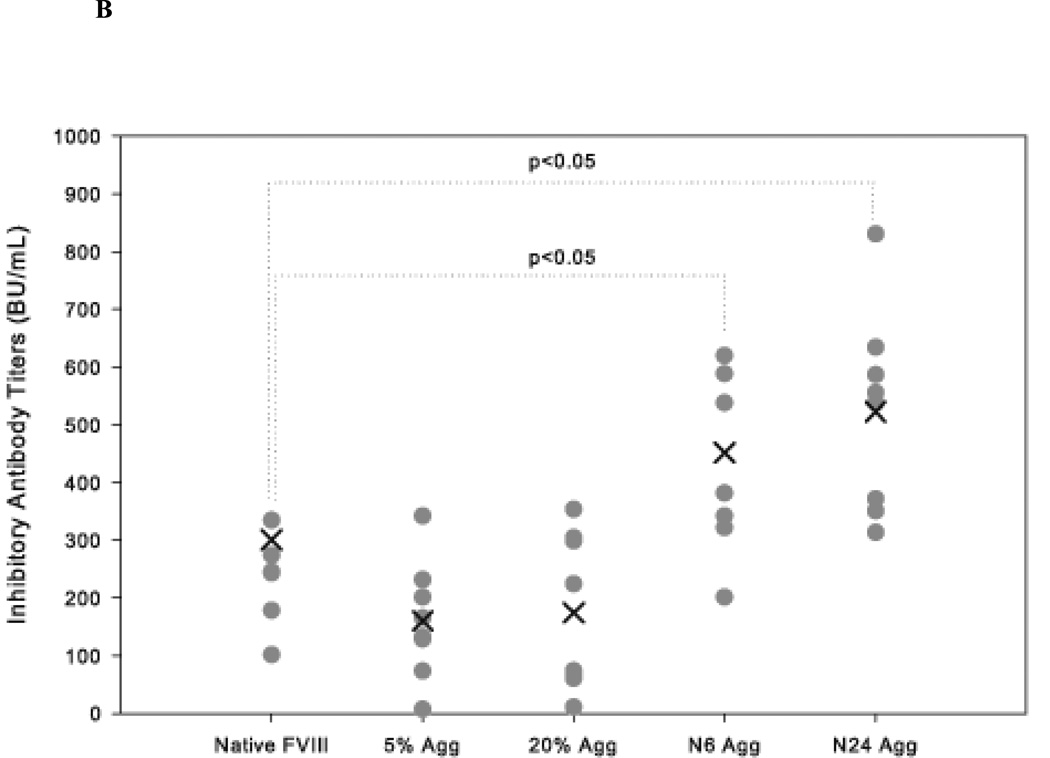
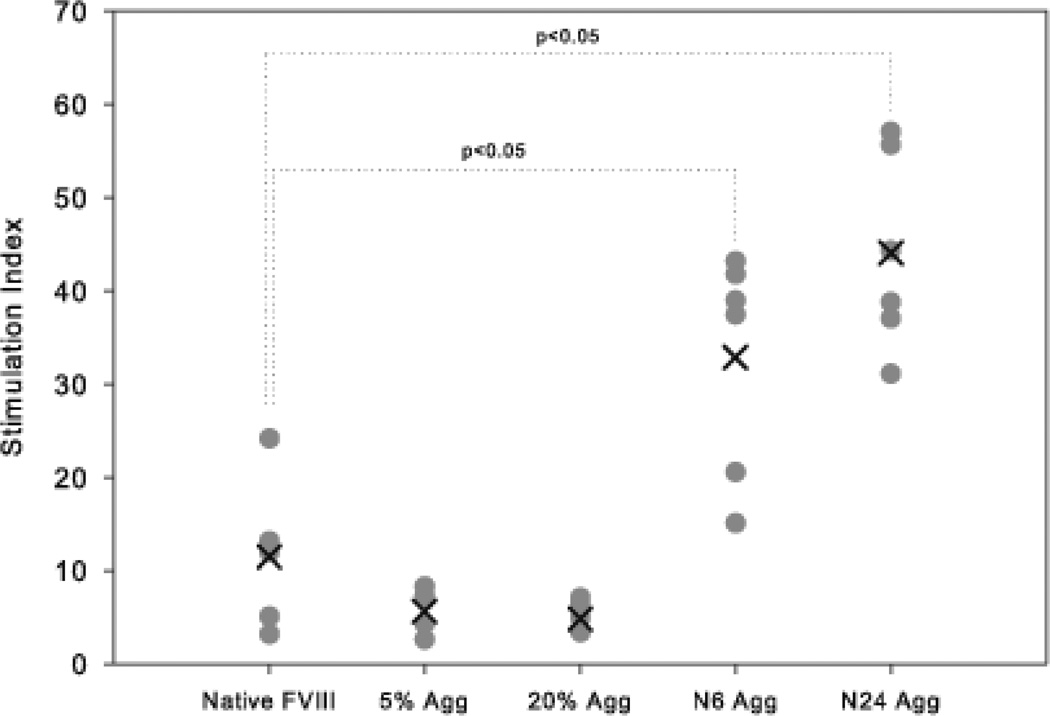
The administration of recombinant factor VIII (FVIII) is the first-line therapy for hemophilia A (HA), but 25%-35% of patients develop an inhibitory antibody response. In general, the presence of aggregates contributes to unwanted immunogenic responses against therapeutic proteins. FVIII has been shown to form both native-like and nonnative aggregates. Previously, we showed that nonnative aggregates of FVIII are less immunogenic than the native protein. Here, we investigated the effect of native-like aggregates of FVIII on immunogenicity in HA and von Willebrand factor knockout (vWF(-/-)) mice. Mice immunized with native-like aggregates showed significantly higher inhibitory antibody titers than animals that received native FVIII. Following restimulation in vitro with native FVIII, the activation of CD4+ T-cells isolated from mice immunized with native-like aggregates is approximately fourfold higher than mice immunized with the native protein. Furthermore, this is associated with increases in the secretion of proinflammatory cytokines IL-6 and IL-17 in the native-like aggregate treatment group. The results indicate that the native-like aggregates of FVIII are more immunogenic than native FVIII for both the B-cell and the T-cell responses.
Copyright © 2012 Wiley Periodicals, Inc.
Figures
Similar articles
-
The important role of von Willebrand factor in platelet-derived FVIII gene therapy for murine hemophilia A in the presence of inhibitory antibodies.J Thromb Haemost. 2015 Jul;13(7):1301-9. doi: 10.1111/jth.13001. Epub 2015 Jun 11. J Thromb Haemost. 2015. PMID: 25955153 Free PMC article.
-
Evaluation of von Willebrand factor phenotypes and genotypes in Hemophilia A patients with and without identified F8 mutations.J Thromb Haemost. 2015 Jun;13(6):1036-42. doi: 10.1111/jth.12902. Epub 2015 May 9. J Thromb Haemost. 2015. PMID: 25780857 Free PMC article.
-
Anti-C1 domain antibodies that accelerate factor VIII clearance contribute to antibody pathogenicity in a murine hemophilia A model.J Thromb Haemost. 2018 Sep;16(9):1779-1788. doi: 10.1111/jth.14233. Epub 2018 Aug 13. J Thromb Haemost. 2018. PMID: 29981270 Free PMC article.
-
To serve and protect: The modulatory role of von Willebrand factor on factor VIII immunogenicity.Blood Rev. 2017 Sep;31(5):339-347. doi: 10.1016/j.blre.2017.07.001. Epub 2017 Jul 4. Blood Rev. 2017. PMID: 28716211 Review.
-
Factor VIII-von Willebrand factor binding defects in autosomal recessive von Willebrand disease type Normandy and in mild hemophilia A. New insights into factor VIII-von Willebrand factor interactions.Acta Haematol. 2009;121(2-3):102-5. doi: 10.1159/000214849. Epub 2009 Jun 8. Acta Haematol. 2009. PMID: 19506355 Review.
Cited by
-
Immunogenicity of Bioproducts: Cellular Models to Evaluate the Impact of Therapeutic Antibody Aggregates.Front Immunol. 2020 May 5;11:725. doi: 10.3389/fimmu.2020.00725. eCollection 2020. Front Immunol. 2020. PMID: 32431697 Free PMC article. Review.
-
Immunogenicity Challenges Associated with Subcutaneous Delivery of Therapeutic Proteins.BioDrugs. 2021 Mar;35(2):125-146. doi: 10.1007/s40259-020-00465-4. Epub 2021 Feb 1. BioDrugs. 2021. PMID: 33523413 Free PMC article. Review.
-
Protein Nanoparticles Promote Microparticle Formation in Intravenous Immunoglobulin Solutions During Freeze-Thawing and Agitation Stresses.J Pharm Sci. 2018 Jul;107(7):1852-1857. doi: 10.1016/j.xphs.2018.03.016. Epub 2018 Mar 27. J Pharm Sci. 2018. PMID: 29601840 Free PMC article.
-
Immunogenicity of subcutaneously administered therapeutic proteins--a mechanistic perspective.AAPS J. 2013 Oct;15(4):897-900. doi: 10.1208/s12248-013-9510-6. Epub 2013 Jul 16. AAPS J. 2013. PMID: 23856740 Free PMC article. Review.
-
Microparticles and Nanoparticles Delivered in Intravenous Saline and in an Intravenous Solution of a Therapeutic Antibody Product.J Pharm Sci. 2017 Feb;106(2):511-520. doi: 10.1016/j.xphs.2016.09.028. Epub 2016 Nov 7. J Pharm Sci. 2017. PMID: 27832839 Free PMC article.
References
-
- Ewenstein BM, Collins P, Tarantino MD, Negrier C, Blanchette V, Shapiro AD, Baker D, Spotts G, Sensel M, Yi SE, Gomperts ED. Hemophilia therapy innovation: development of an advanced category recombinant factor VIII by a plasma/albumin-free method. Proceedings of a Special Symposium at the XIXth Congress of the International Society on Thrombosis and Haemostasis; July 12–18, 2003; Semin Hematol; Birmingham, UK. pp. 1–16. discussion 16–18. - PubMed
-
- Kempton CL, White GC., 2nd. How we treat a hemophilia A patient with a factor VIII inhibitor. Blood. 2009;113(1):11–17. - PubMed
-
- Gouw SC, van der Bom JG, Auerswald G, Ettinghausen CE, Tedgard U, van den Berg HM. Recombinant versus plasma-derived factor VIII products and the development of inhibitors in previously untreated patients with severe hemophilia A: the CANAL cohort study. Blood. 2007;109(11):4693–4697. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
