

Recruitment of coregulator G9a by Runx2 for selective enhancement or suppression of transcription
- PMID: 22389001
- PMCID: PMC3350606
- DOI: 10.1002/jcb.24114
Recruitment of coregulator G9a by Runx2 for selective enhancement or suppression of transcription
Abstract
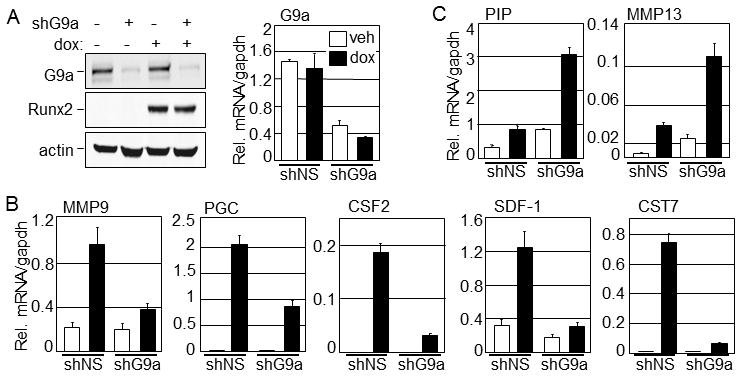
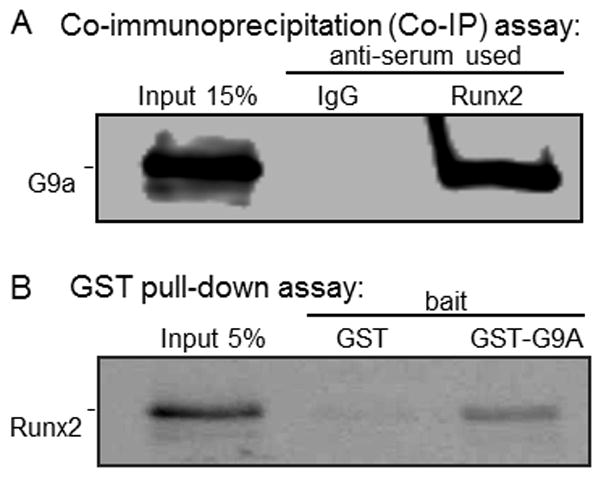
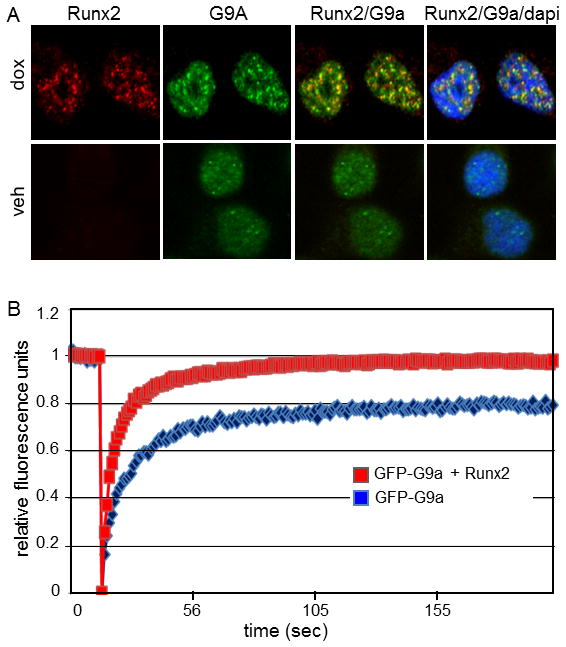
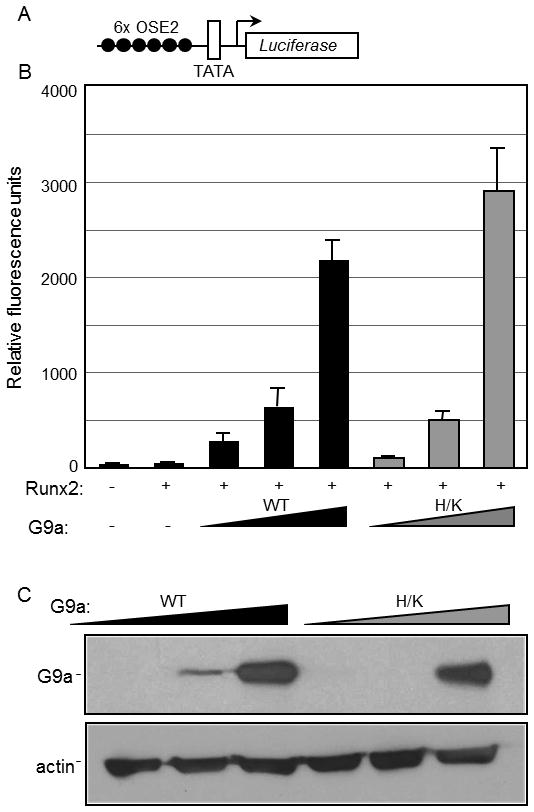
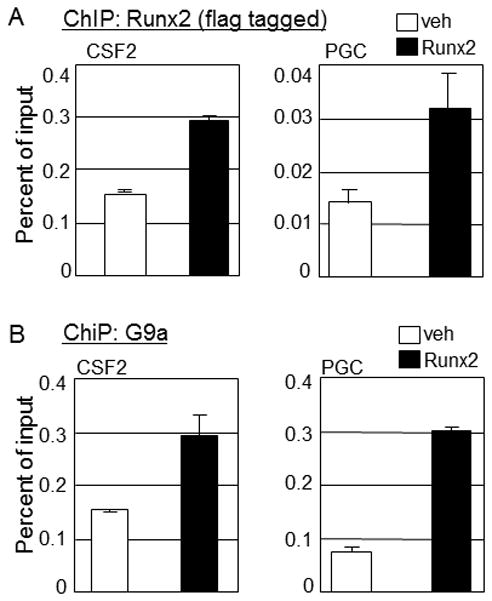
Runx2, best known for its role in regulating osteoblast-specific gene expression, also plays an increasingly recognized role in prostate and breast cancer metastasis. Using the C4-2B/Rx2(dox) prostate cancer cell line that conditionally expressed Runx2 in response to doxycycline treatment, we identified and characterized G9a, a histone methyltransferase, as a novel regulator for Runx2 activity. G9a function was locus-dependent. Whereas depletion of G9a reduced expression of many Runx2 target genes, including MMP9, CSF2, SDF1, and CST7, expression of others, such as MMP13 and PIP, was enhanced. Physical association between G9a and Runx2 was indicated by co-immunoprecipitation, GST-pulldown, immunofluorescence, and fluorescence recovery after photobleaching (FRAP) assays. Since G9a makes repressive histone methylation marks and is primarily known as a corepressor, we further investigated the mechanism by which G9a functioned as a positive regulator for Runx2 target genes. Transient reporter assays indicated that the histone methyltransferase activity of G9a was not required for transcriptional activation by Runx2. Chromatin immunoprecipitation assays for Runx2 and G9a showed that G9a was recruited to endogenous Runx2 binding sites. We conclude that a subset of cancer-related Runx2 target genes require recruitment of G9a for their expression, but do not depend on its histone methyltransferase activity.
Copyright © 2012 Wiley Periodicals, Inc.
Conflict of interest statement
All authors declare no conflict of interest.
Figures
References
-
- Akech J, Wixted JJ, Bedard K, van der Deen M, Hussain S, Guise TA, van Wijnen AJ, Stein JL, Languino LR, Altieri DC, Pratap J, Keller E, Stein GS, Lian JB. Runx2 association with progression of prostate cancer in patients: mechanisms mediating bone osteolysis and osteoblastic metastatic lesions. Oncogene. 2010;29:811–21. - PMC - PubMed
-
- Barnes GL, Hebert KE, Kamal M, Javed A, Einhorn TA, Lian JB, Stein GS, Gerstenfeld LC. Fidelity of Runx2 activity in breast cancer cells is required for the generation of metastases-associated osteolytic disease. Cancer Res. 2004;64:4506–13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
