

Phylogenetic framework and molecular signatures for the main clades of the phylum Actinobacteria
- PMID: 22390973
- PMCID: PMC3294427
- DOI: 10.1128/MMBR.05011-11
Phylogenetic framework and molecular signatures for the main clades of the phylum Actinobacteria
Abstract
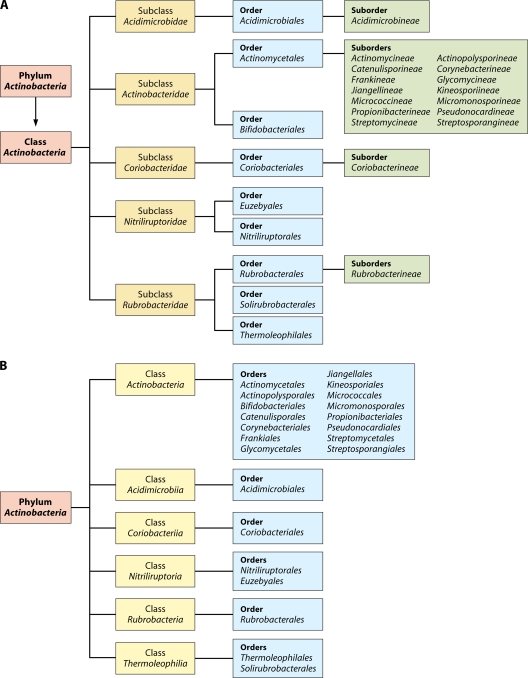
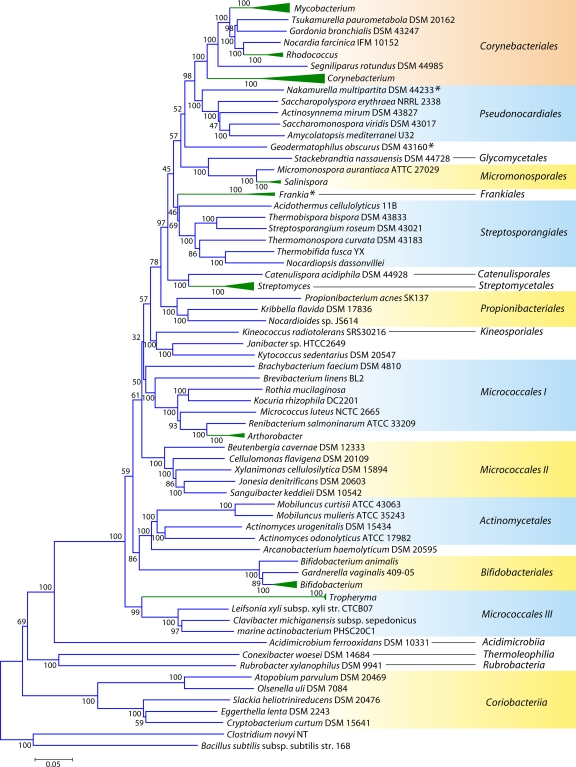
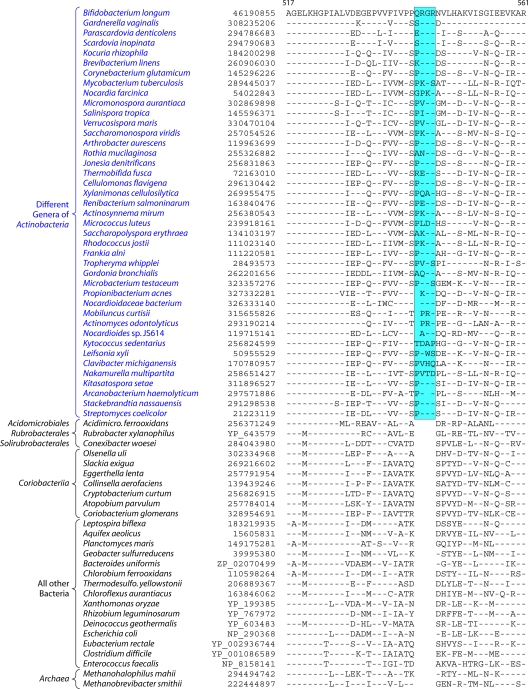
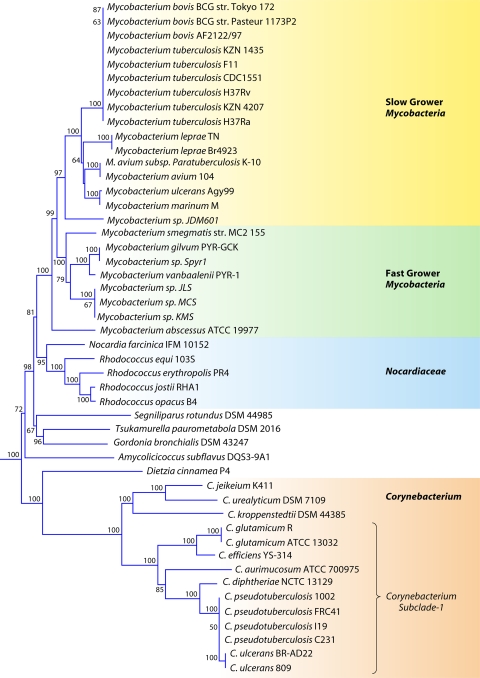
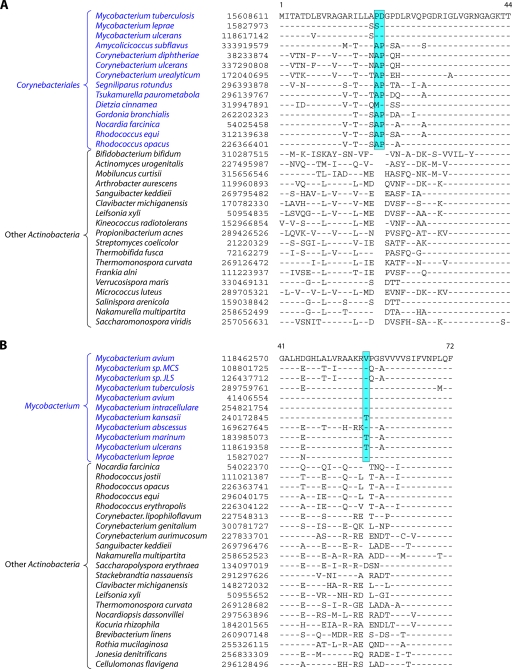
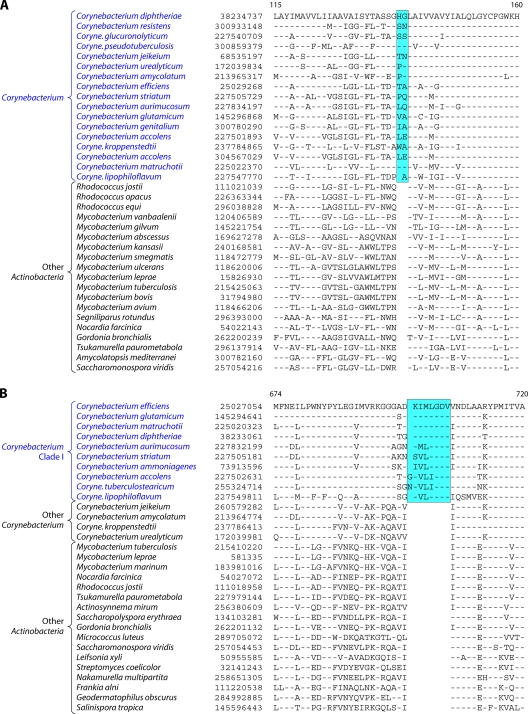
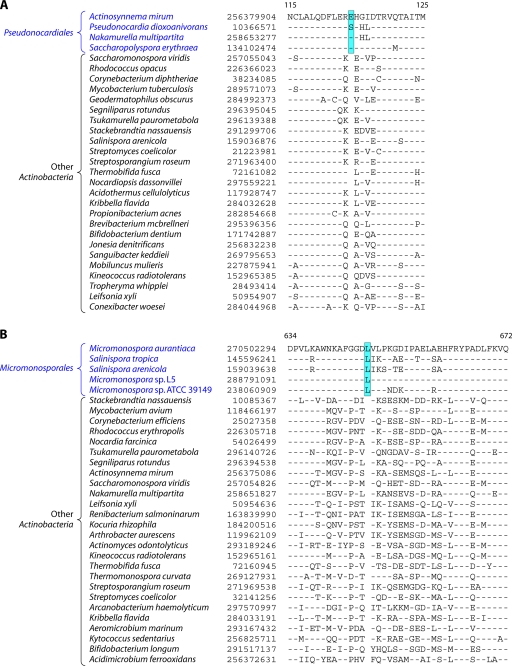
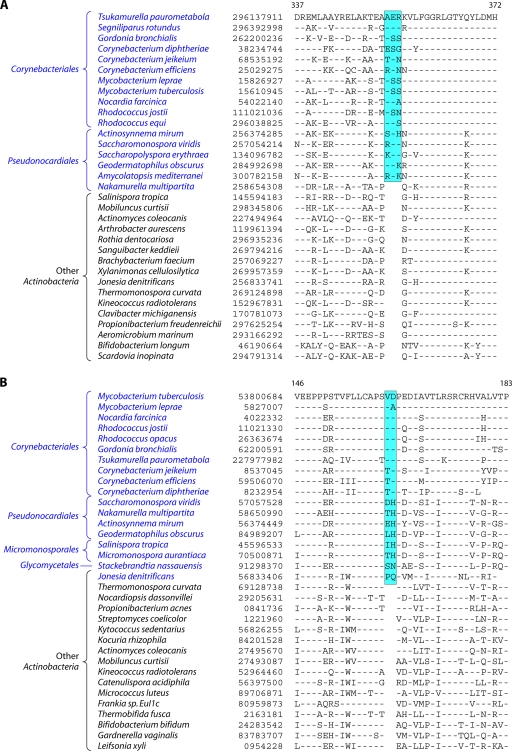
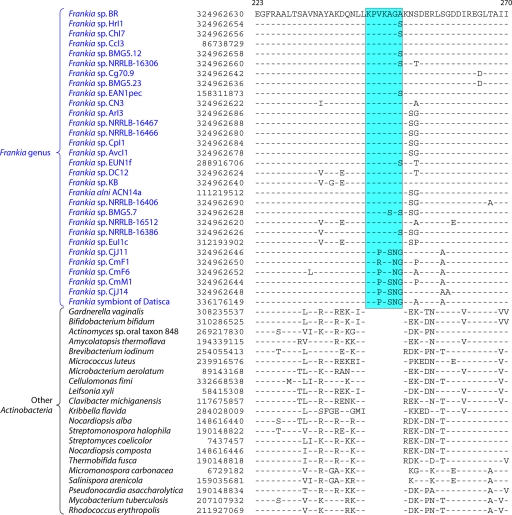
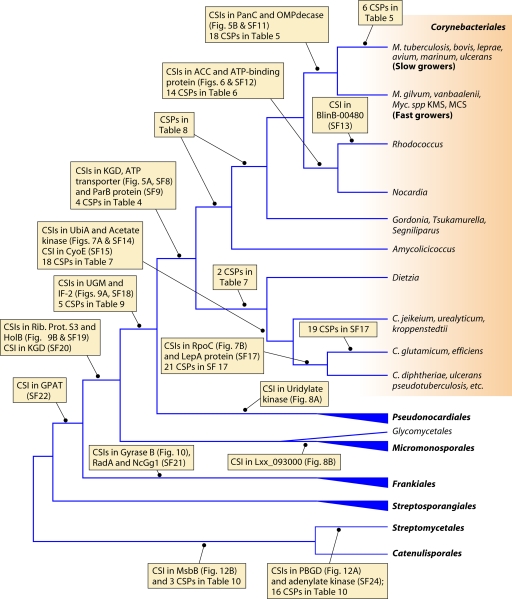
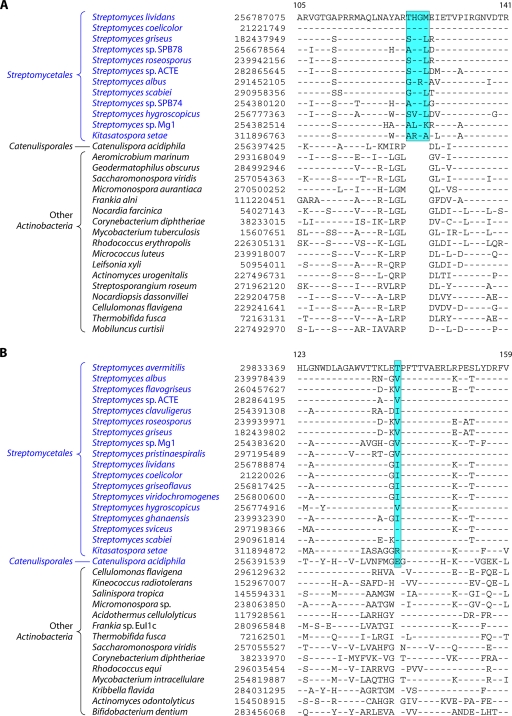
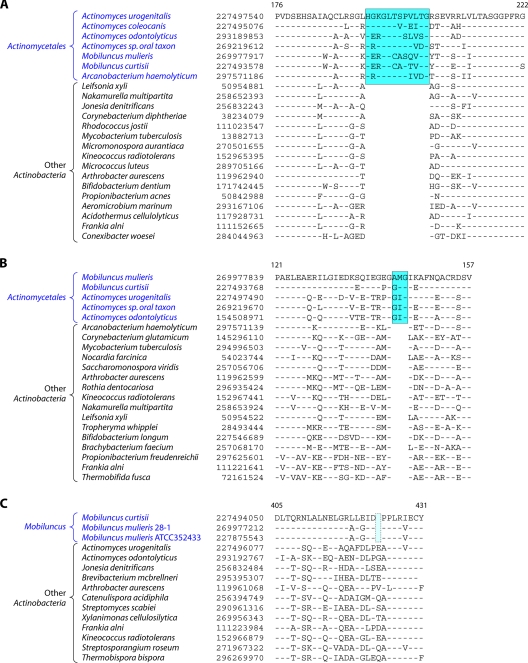
The phylum Actinobacteria harbors many important human pathogens and also provides one of the richest sources of natural products, including numerous antibiotics and other compounds of biotechnological interest. Thus, a reliable phylogeny of this large phylum and the means to accurately identify its different constituent groups are of much interest. Detailed phylogenetic and comparative analyses of >150 actinobacterial genomes reported here form the basis for achieving these objectives. In phylogenetic trees based upon 35 conserved proteins, most of the main groups of Actinobacteria as well as a number of their superageneric clades are resolved. We also describe large numbers of molecular markers consisting of conserved signature indels in protein sequences and whole proteins that are specific for either all Actinobacteria or their different clades (viz., orders, families, genera, and subgenera) at various taxonomic levels. These signatures independently support the existence of different phylogenetic clades, and based upon them, it is now possible to delimit the phylum Actinobacteria (excluding Coriobacteriia) and most of its major groups in clear molecular terms. The species distribution patterns of these markers also provide important information regarding the interrelationships among different main orders of Actinobacteria. The identified molecular markers, in addition to enabling the development of a stable and reliable phylogenetic framework for this phylum, also provide novel and powerful means for the identification of different groups of Actinobacteria in diverse environments. Genetic and biochemical studies on these Actinobacteria-specific markers should lead to the discovery of novel biochemical and/or other properties that are unique to different groups of Actinobacteria.
Figures


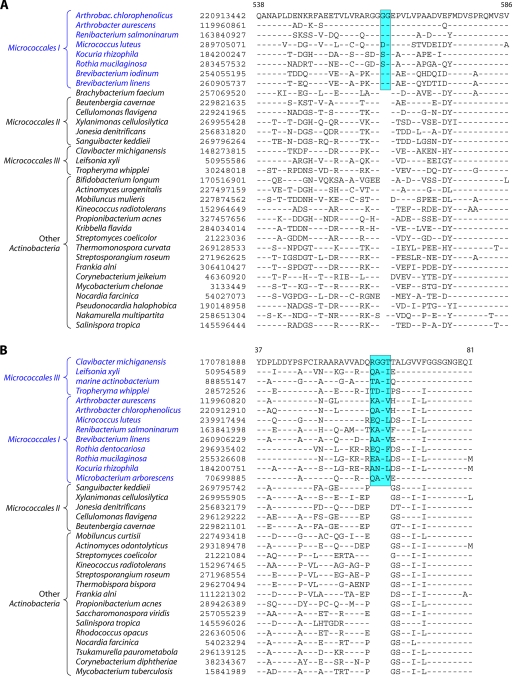

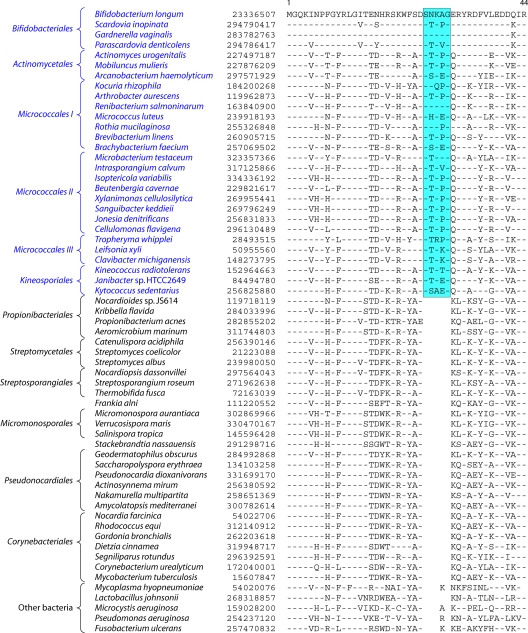
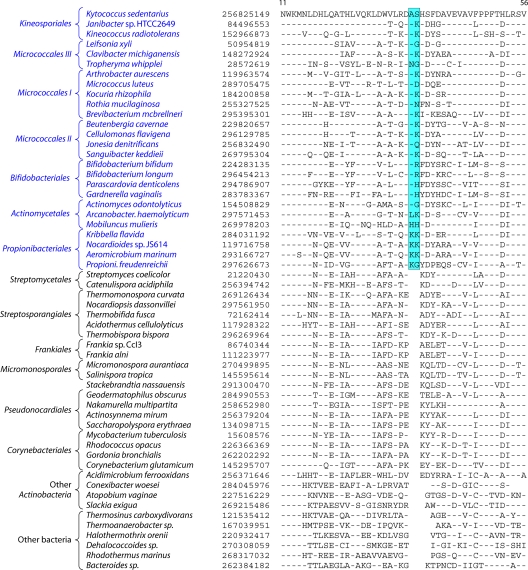
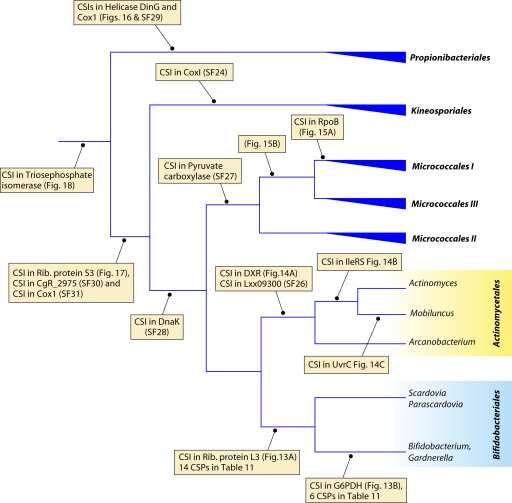


References
-
- Abdallah AM, et al. 2006. A specific secretion system mediates PPE41 transport in pathogenic mycobacteria. Mol. Microbiol. 62:667–679 - PubMed
-
- Alam MT, Merlo ME, Takano E, Breitling R. 2010. Genome-based phylogenetic analysis of Streptomyces and its relatives. Mol. Phylogenet. Evol. 54:763–772 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
