

Review
doi: 10.1083/jcb.201105099.
The cell biology of regeneration
Affiliations
- PMID: 22391035
- PMCID: PMC3307701
- DOI: 10.1083/jcb.201105099
Item in Clipboard
Review
The cell biology of regeneration
J Cell Biol.
.
Abstract
Regeneration of complex structures after injury requires dramatic changes in cellular behavior. Regenerating tissues initiate a program that includes diverse processes such as wound healing, cell death, dedifferentiation, and stem (or progenitor) cell proliferation; furthermore, newly regenerated tissues must integrate polarity and positional identity cues with preexisting body structures. Gene knockdown approaches and transgenesis-based lineage and functional analyses have been instrumental in deciphering various aspects of regenerative processes in diverse animal models for studying regeneration.
Figures
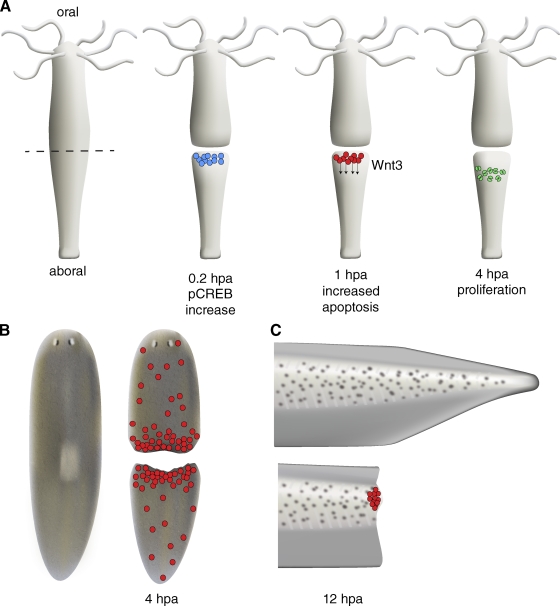
Increased apoptosis is associated with early phases of regeneration. (A) After mid-gastric bisection in Hydra, MAPK signaling leads to rapid activation of the transcription factor CREB in fragments regenerating a head (blue cells). MAPK/CREB activity is required for stimulating a wave of apoptosis in interstitial cells near the site of injury (red cells). These apoptotic cells secrete Wnt3, inducing a zone of proliferation (green cells) below the region of apoptosis. (B) Planarians and (C) Xenopus larval tails also show a rapid, localized increase in apoptosis after amputation. In Xenopus, like Hydra, apoptosis may provide important signals during early phases of regeneration; inhibiting apoptosis during the first 24 h post amputation (hpa) blocks regeneration.
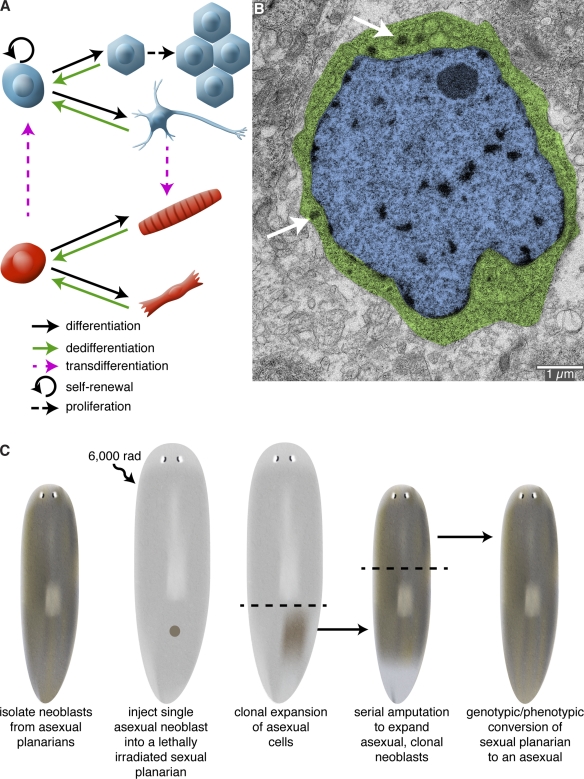
Cellular sources of regeneration. (A) The ability to regenerate amputated structures often requires the production of new cells. These new cells can be derived from amplification and differentiation of resident stem cells, proliferation of differentiated cells, dedifferentiation of cells to a more primitive state, or transdifferentiation of one cell type to another cell type. (B) Transmission electron micrograph of a neoblast from the planarian Schmidtea mediterranea. Planarians owe their impressive regenerative abilities to these adult stem cells. Neoblasts are the only mitotic somatic cells and are defined by their high nuclear (blue) to cytoplasm (green) ratio and the presence of cytoplasmic ribonucleoprotein complexes called chromatoid bodies (arrows). Image courtesy of Ana Vieira (University of Illinois at Urbana-Champaign, Urbana, IL). (C) Rescue of a lethally irradiated planarian by introduction of a single neoblast. S. mediterranea exists as two genetically distinct strains: an asexual strain (brown) that reproduces by transverse fission; and a sexual, hermaphroditic strain (gray) that reproduces by cross-fertilization. Wagner et al. (2011) have shown that lethally irradiated sexual animals can be rescued by injection of a single asexual neoblast. Because the asexual donor neoblast is the only source of new cells, the sexual host is eventually converted into an asexual animal after repeated rounds of amputation, regeneration, and tissue turnover.
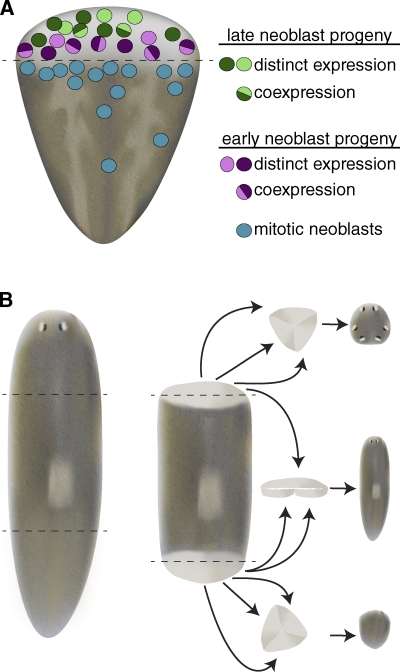
Neoblast progeny are specified during early phases of regeneration yet retain developmental plasticity. (A) The planarian blastema is composed of a heterogeneous mixture of differentiating cells. The blastema is largely devoid of proliferating neoblasts, and instead is composed of cells expressing early and late neoblast progeny markers with distinct spatial distributions. Although some cells coexpress early (or late) neoblast progeny markers, other cells show distinct expression profiles for various early neoblast progeny markers, suggesting early neoblast progeny may be en route to committing to terminal cell types (adapted from Eisenhoffer et al., 2008). (B) 3-d anterior blastemas (top) are capable of forming anterior structures, including photoreceptors and cephalic ganglia, after being surgically isolated from intact tissue and cultured in vitro. Similarly, 3-d posterior blastemas (bottom) repigment and form muscle, but do not generate anterior structures, suggesting that by 3 d of regeneration head or tail specification has occurred. When 3-d anterior and posterior blastemas are juxtaposed (middle) they can generate a planarian containing mid-body structures (including a pharynx), in addition to head and tail tissue. These results indicate that at least some of the post-mitotic neoblast progeny that compose the blastema are capable of altering their fates (adapted from Sengel, 1960).
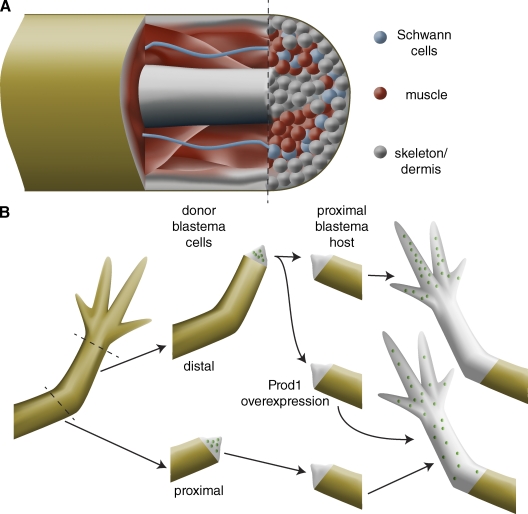
Cellular memory during vertebrate limb regeneration. (A) Although the regeneration blastema appears to be a homogeneous mass of undifferentiated cells, lineage-tracing experiments in the axolotl limb, the Xenopus tail, and the zebrafish caudal fin indicate that blastema cells only contribute to tissues of similar developmental origin as that from which they are derived. For example, muscle (red) only gives rise to new muscle. Although dermis (gray) can give rise to new skeletal elements, these tissues are both lateral plate mesoderm derivatives, suggesting only limited dedifferentiation. Therefore, the blastema is composed of a heterogeneous mixture of lineage-restricted cells and is not a homogeneous population of multipotent cells. (B) Blastema cells retain their proximo-distal identity: distal amputations produce blastemas that only regenerate distal structures, whereas more proximal amputations produce blastemas that regenerate medial and distal structures. When cells from a distal blastema (green dots) are transplanted into a proximal blastema, they contribute only to distal structures; by contrast, cells from a proximal blastema (green dots) contribute to structures along the length of the proximo-distal axis. Overexpression of the cell surface protein Prod1 transforms distal blastema cells to more proximal fates.
References
-
- Baguñà J., Saló E., Auladell C. 1989. Regeneration and pattern formation in planarians III. Evidence that neoblasts are totipotent stem cells and the source of blastema cells. Development. 107:77–86
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
