

NLRP3 inflammasome: key mediator of neuroinflammation in murine Japanese encephalitis
- PMID: 22393394
- PMCID: PMC3290554
- DOI: 10.1371/journal.pone.0032270
NLRP3 inflammasome: key mediator of neuroinflammation in murine Japanese encephalitis
Abstract
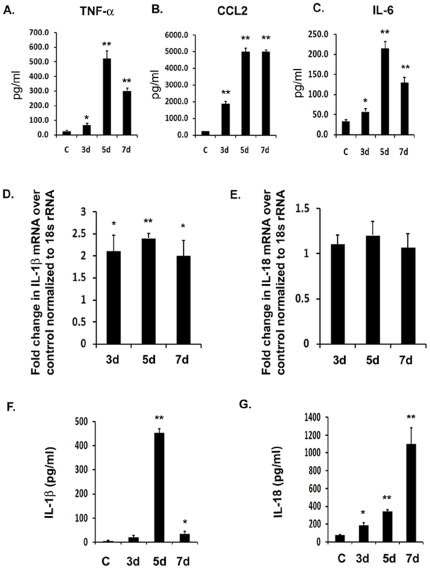
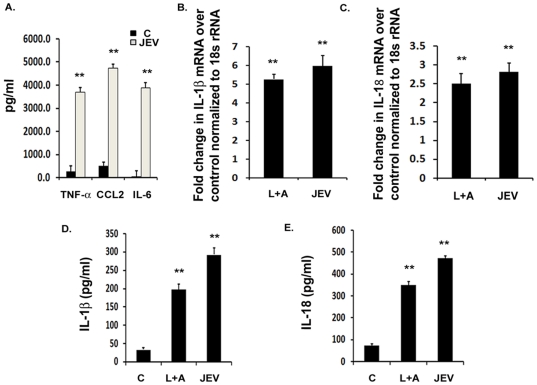
Background: Japanese Encephalitis virus (JEV) is a common cause of acute and epidemic viral encephalitis. JEV infection is associated with microglial activation resulting in the production of pro-inflammatory cytokines including Interleukin-1 β (IL-1β) and Interleukin-18 (IL-18). The Pattern Recognition Receptors (PRRs) and the underlying mechanism by which microglia identify the viral particle leading to the production of these cytokines is unknown.
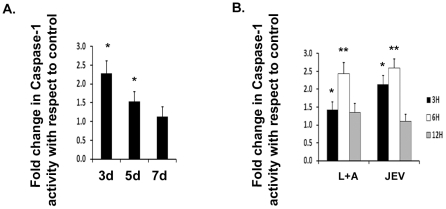
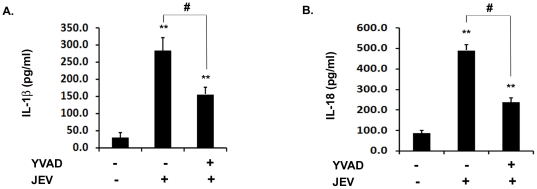
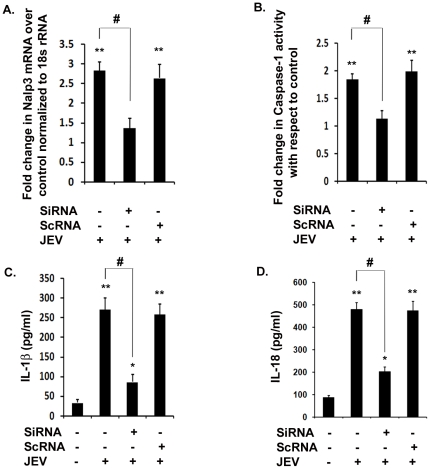
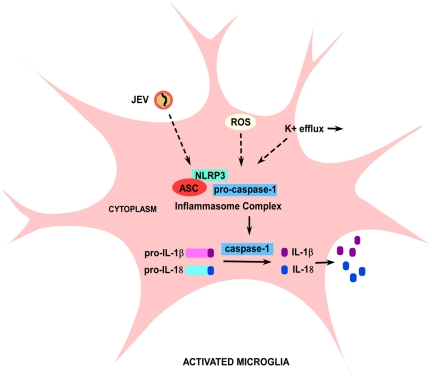
Methodology/principal findings: For our studies, we have used murine model of JEV infection as well as BV-2 mouse microglia cell line. In this study, we have identified a signalling pathway which leads to the activation of caspase-1 as the key enzyme responsible for the maturation of both IL-1β and IL-18 in NACHT, LRR and PYD domains-containing protein-3 (NLRP3) dependent manner. Depletion of NLRP3 results in the reduction of caspase-1 activity and subsequent production of these cytokines.
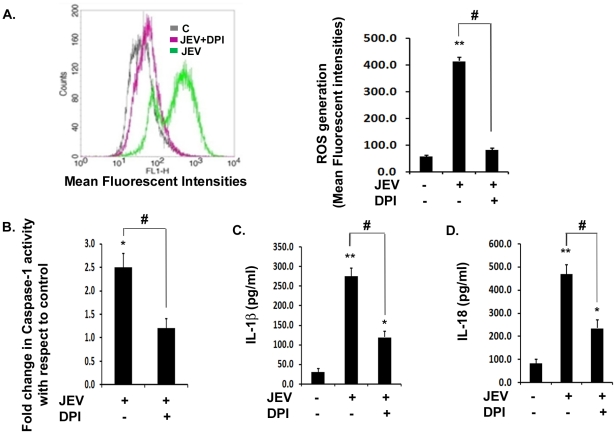
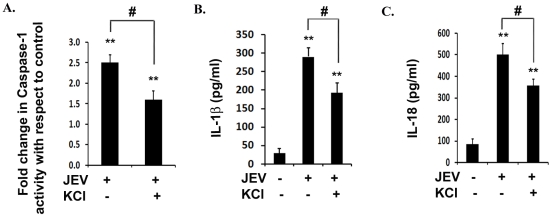
Conclusion/significance: Our results identify a mechanism mediated by Reactive Oxygen Species (ROS) production and potassium efflux as the two danger signals that link JEV infection to caspase-1 activation resulting in subsequent IL-1β and IL-18 maturation.
Conflict of interest statement
Figures
References
-
- Krady JK, Basu A, Levison SW, Milner RJ. Differential expression of protein tyrosine kinase genes during microglial activation. Glia. 2002;40:11–24. - PubMed
-
- Levison SW, Druckman SK, Young GM, Basu A. Neural stem cells in the subventricular zone are a source of astrocytes and oligodendrocytes, but not microglia. Dev Neurosci. 2003;25:184–196. - PubMed
-
- Kaushik DK, Gupta M, Basu A. Microglial response to viral challenges: Every silver lining comes with a cloud. Front Biosci. 2011;17:2187–2205. - PubMed
-
- Westaway EG, Brinton MA, Gaidamovich S, Horzinek MC, Igarashi A, et al. Flaviviridae. Intervirology. 1985;24:183–192. - PubMed
-
- Kumar R, Mathur A, Kumar A, Sharma S, Chakraborty S, et al. Clinical features & prognostic indicators of Japanese encephalitis in children in Lucknow (India). Indian J Med Res. 1990;91:321–327. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
