

Roles of the amino terminal region and repeat region of the Plasmodium berghei circumsporozoite protein in parasite infectivity
- PMID: 22393411
- PMCID: PMC3290588
- DOI: 10.1371/journal.pone.0032524
Roles of the amino terminal region and repeat region of the Plasmodium berghei circumsporozoite protein in parasite infectivity
Abstract
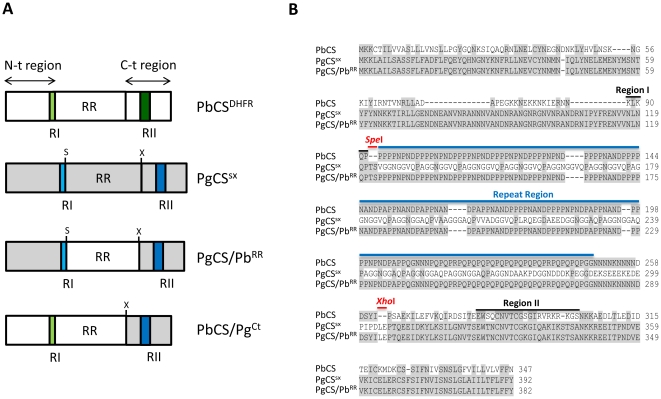
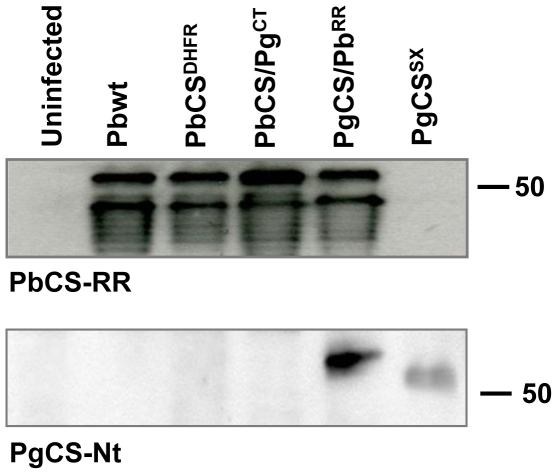
The circumsporozoite protein (CSP) plays a key role in malaria sporozoite infection of both mosquito salivary glands and the vertebrate host. The conserved Regions I and II have been well studied but little is known about the immunogenic central repeat region and the N-terminal region of the protein. Rodent malaria Plasmodium berghei parasites, in which the endogenous CS gene has been replaced with the avian Plasmodium gallinaceum CS (PgCS) sequence, develop normally in the A. stephensi mosquito midgut but the sporozoites are not infectious. We therefore generated P. berghei transgenic parasites carrying the PgCS gene, in which the repeat region was replaced with the homologous region of P. berghei CS (PbCS). A further line, in which both the N-terminal region and repeat region were replaced with the homologous regions of PbCS, was also generated. Introduction of the PbCS repeat region alone, into the PgCS gene, did not rescue sporozoite species-specific infectivity. However, the introduction of both the PbCS repeat region and the N-terminal region into the PgCS gene completely rescued infectivity, in both the mosquito vector and the mammalian host. Immunofluorescence experiments and western blot analysis revealed correct localization and proteolytic processing of CSP in the chimeric parasites. The results demonstrate, in vivo, that the repeat region of P. berghei CSP, alone, is unable to mediate sporozoite infectivity in either the mosquito or the mammalian host, but suggest an important role for the N-terminal region in sporozoite host cell invasion.
Conflict of interest statement
Figures
References
-
- Nardin E, Zavala F, Nussenzweig V, Nussenzweig RS. Pre-erythrocytic malaria vaccine: mechanisms of protective immunity and human vaccine trials. Parassitologia. 1999;41:397–402. - PubMed
-
- Birkett A, Lyons K, Schmidt A, Boyd D, Oliveira GA, et al. A modified hepatitis B virus core particle containing multiple epitopes of the Plasmodium falciparum circumsporozoite protein provides a highly immunogenic malaria vaccine in preclinical analyses in rodent and primate hosts. Infect Immun. 2002;70:6860–6870. - PMC - PubMed
-
- Ballou WR, Hoffman SL, Sherwood JA, Hollingdale MR, Neva FA, et al. Safety and efficacy of a recombinant DNA Plasmodium falciparum sporozoite vaccine. Lancet. 1987;1:1277–1281. - PubMed
-
- Herrington DA, Clyde DF, Losonsky G, Cortesia M, Murphy JR, et al. Safety and immunogenicity in man of a synthetic peptide malaria vaccine against Plasmodium falciparum sporozoites. Nature. 1987;328:257–259. - PubMed
-
- Ballou WR. The development of the RTS,S malaria vaccine candidate: challenges and lessons. Parasite Immunol. 2009;31:492–500. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
