

In-depth characterization of viral isolates from plasma and cells compared with plasma circulating quasispecies in early HIV-1 infection
- PMID: 22393441
- PMCID: PMC3290612
- DOI: 10.1371/journal.pone.0032714
In-depth characterization of viral isolates from plasma and cells compared with plasma circulating quasispecies in early HIV-1 infection
Abstract
Background: The use of in vitro models to unravel the phenotypic characteristics of circulating viral variants is key to understanding HIV-1 pathogenesis but limited by the availability of primary viral isolates from biological samples. However, overall in vivo genetic variability of HIV-1 within a subject may not be reflected in the viable viral population obtained after isolation. Although several studies have tried to determine whether viral populations expanded in vitro are representative of in vivo findings, the answer remains unclear due to the reduced number of clonal sequences analyzed or samples compared. In order to overcome previous experimental limitations, here we applied Deep Pyrosequencing (DPS) technology in combination with phenotypic experiments to analyze and compare with unprecedented detail the composition of viral isolates and in vivo quasispecies.
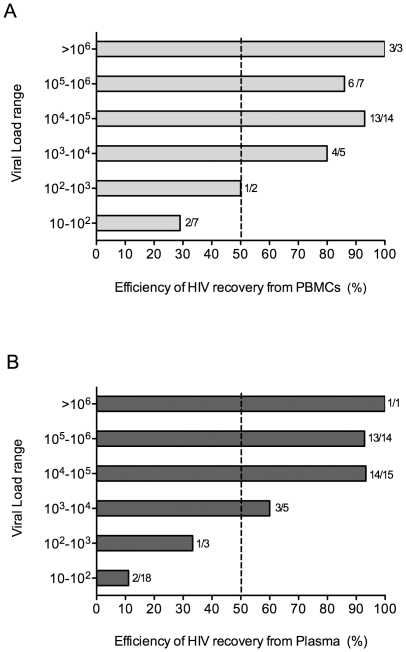
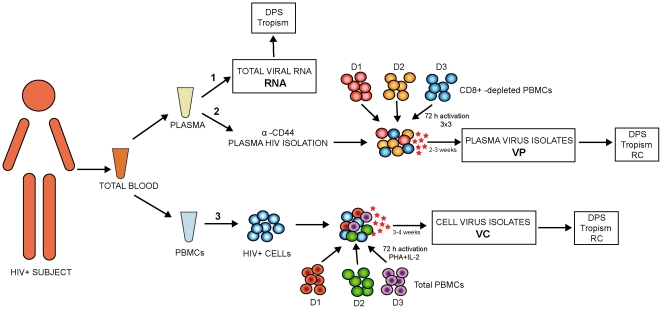
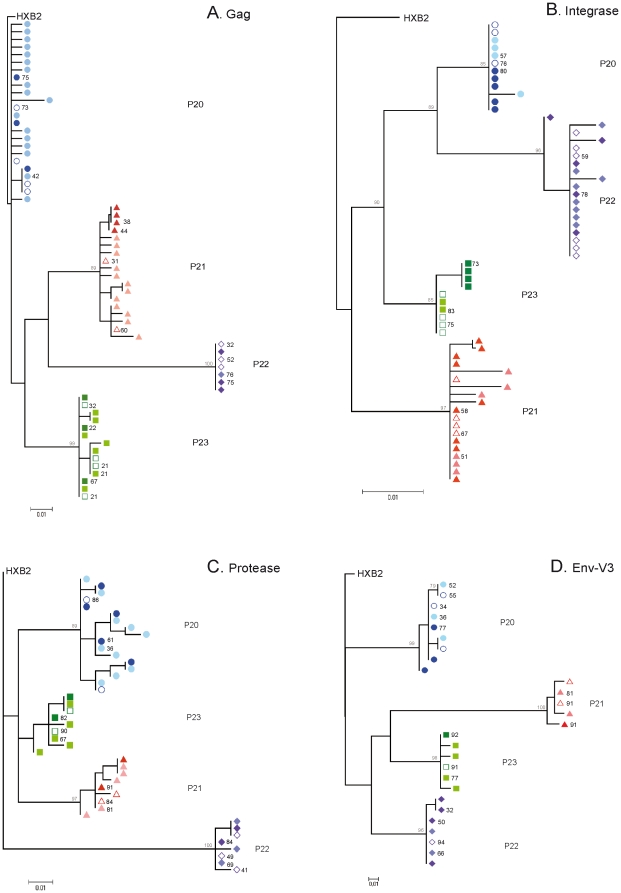
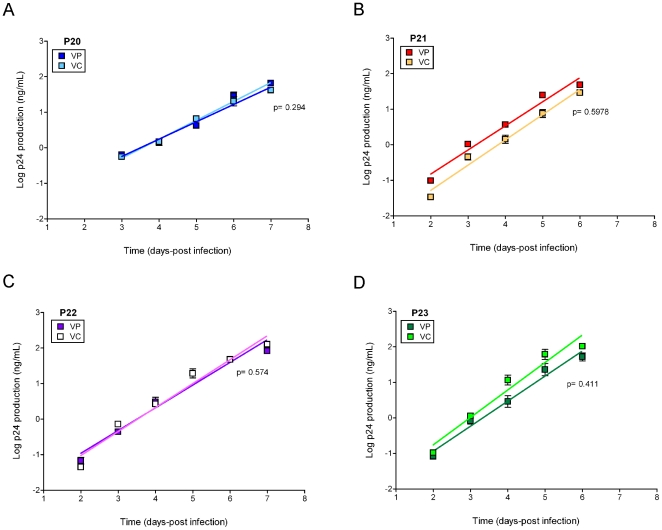
Methodology/principal findings: We amplified by DPS HIV-1 genomic regions covering gag, protease, integrase and env-V3 to characterize paired isolates from plasma and peripheral blood mononuclear cells and compare them with total plasma viral RNA in four recently HIV-1 infected subjects. Our study demonstrated the presence of unique haplotypes scattered between sample types with conservation of major variants. In addition, no differences in intra- and inter-population encoded protein variability were found between the different types of isolates or when these were compared to plasma viral RNA within subjects. Additionally, in vitro experiments demonstrated phenotypic similarities in terms of replicative capacity and co-receptor usage between viral isolates and plasma viral RNA.
Conclusion: This study is the first in-depth comparison and characterization of viral isolates from different sources and plasma circulating quasispecies using DPS in recently HIV-1 infected subjects. Our data supports the use of primary isolates regardless of their plasma or cellular origin to define genetic variability and biological traits of circulating HIV-1 quasispecies.
Conflict of interest statement
Figures
References
-
- Harris RS, Bishop KN, Sheehy AM, Craig HM, Petersen-Mahrt SK, et al. DNA deamination mediates innate immunity to retroviral infection. Cell. 2003;113:803–809. - PubMed
-
- Ho DD, Neumann AU, Perelson AS, Chen W, Leonard JM, et al. Rapid turnover of plasma virions and CD4 lymphocytes in HIV-1 infection. Nature. 1995;373:123–126. - PubMed
-
- Mangeat B, Turelli P, Caron G, Friedli M, Perrin L, et al. Broad antiretroviral defence by human APOBEC3G through lethal editing of nascent reverse transcripts. Nature. 2003;424:99–103. - PubMed
-
- Perelson AS, Neumann AU, Markowitz M, Leonard JM, Ho DD. HIV-1 dynamics in vivo: virion clearance rate, infected cell life-span, and viral generation time. Science. 1996;271:1582–1586. - PubMed
-
- Preston BD, Poiesz BJ, Loeb LA. Fidelity of HIV-1 reverse transcriptase. Science. 1988;242:1168–1171. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
