

Structural and biochemical characterization of the childhood cataract-associated R76S mutant of human γD-crystallin
- PMID: 22394327
- PMCID: PMC3326406
- DOI: 10.1021/bi300199d
Structural and biochemical characterization of the childhood cataract-associated R76S mutant of human γD-crystallin
Abstract
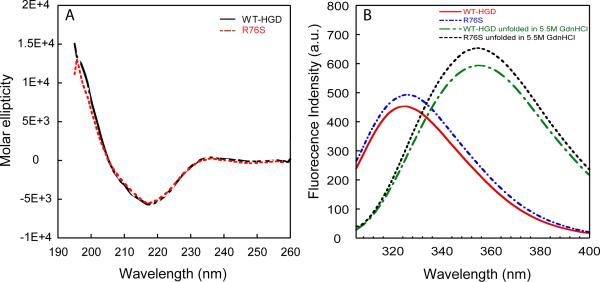
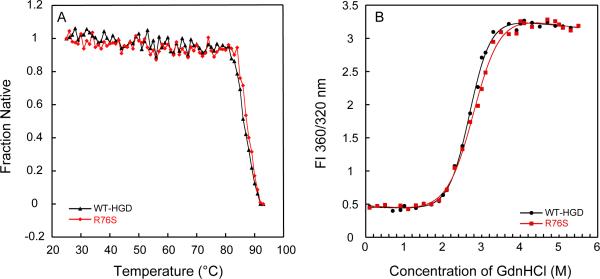
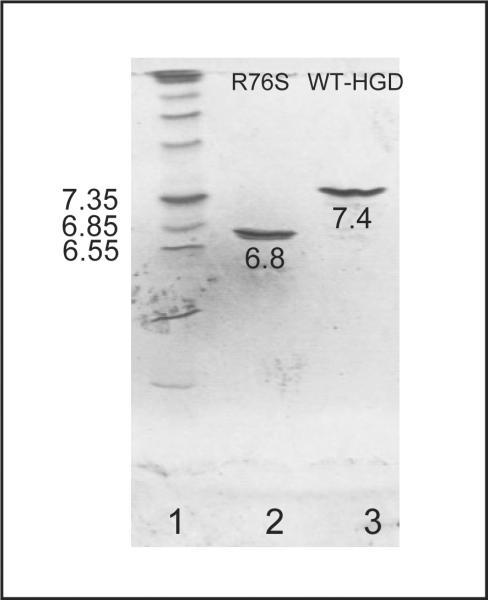
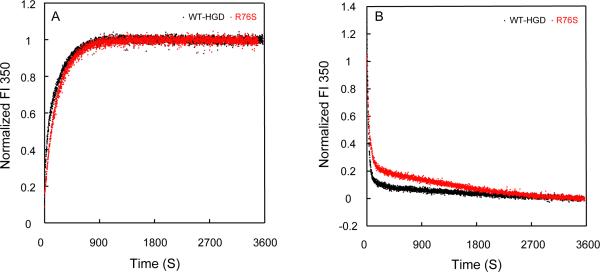
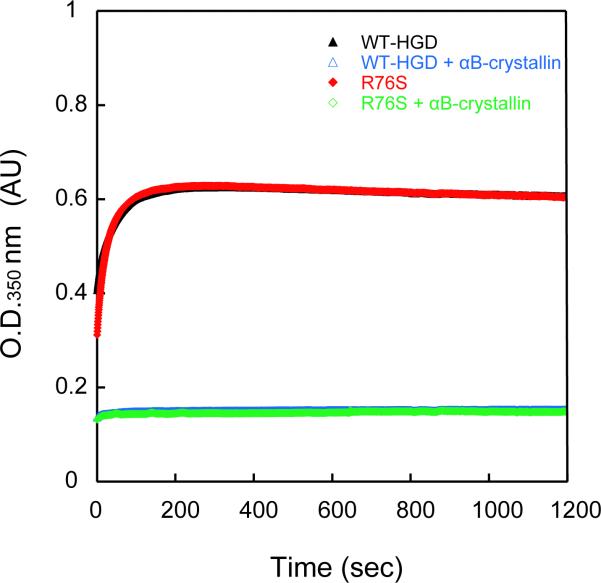
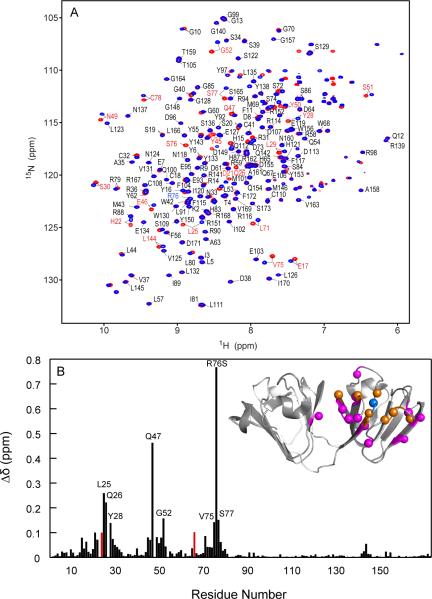
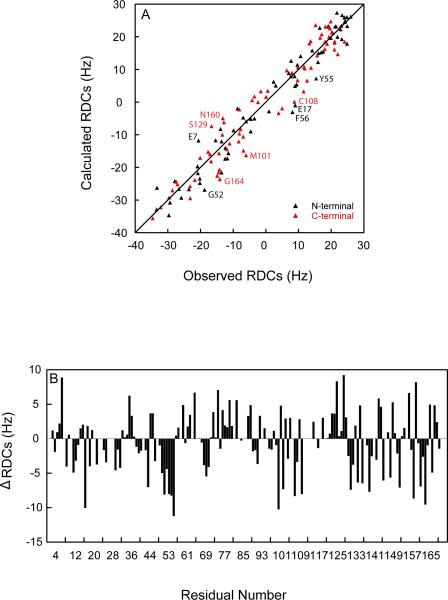
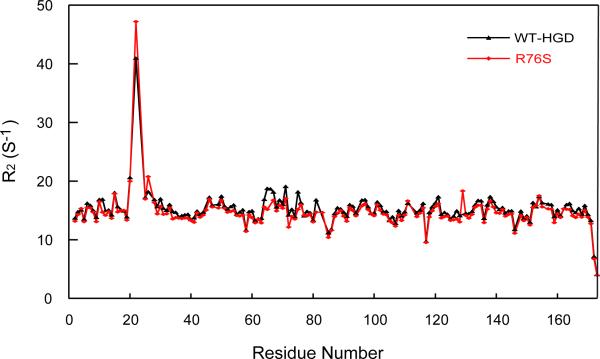
Although a number of γD-crystallin mutations are associated with cataract formation, there is not a clear understanding of the molecular mechanism(s) that lead to this protein deposition disease. As part of our ongoing studies on crystallins, we investigated the recently discovered Arg76 to Ser (R76S) mutation that is correlated with childhood cataract in an Indian family. We expressed the R76S γD-crystallin protein in E. coli, characterized it by CD, fluorescence, and NMR spectroscopy, and determined its stability with respect to thermal and chemical denaturation. Surprisingly, no significant biochemical or biophysical differences were observed between the wild-type protein and the R76S variant, except a lowered pI (6.8 compared to the wild-type value of 7.4). NMR assessment of the R76S γD-crystallin solution structure, by RDCs, and of its motional properties, by relaxation measurements, also revealed a close resemblance to wild-type crystallin. Further, kinetic unfolding/refolding experiments for R76S and wild-type protein showed similar degrees of off-pathway aggregation suppression by αB-crystallin. Overall, our results suggest that neither structural nor stability changes in the protein are responsible for the R76S γD-crystallin variant's association with cataract. However, the change in pI and the associated surface charge or the altered nature of the amino acid could influence interactions with other lens protein species.
Figures
References
-
- Truscott RJ, Augusteyn RC. Oxidative changes in human lens proteins during senile nuclear cataract formation. Biochim Biophys Acta. 1977;492:43–52. - PubMed
-
- Truscott RJ, Augusteyn RC. The state of sulphydryl groups in normal and cataractous human lenses. Exp Eye Res. 1977;25:139–148. - PubMed
-
- Hains PG, Truscott RJ. Post-translational modifications in the nuclear region of young, aged, and cataract human lenses. J Proteome Res. 2007;6:3935–3943. - PubMed
-
- Ma Z, Hanson SR, Lampi KJ, David LL, Smith DL, Smith JB. Age-related changes in human lens crystallins identified by HPLC and mass spectrometry. Exp Eye Res. 1998;67:21–30. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
