

Rosiglitazone selectively inhibits K(ATP) channels by acting on the K(IR) 6 subunit
- PMID: 22394376
- PMCID: PMC3448911
- DOI: 10.1111/j.1476-5381.2012.01934.x
Rosiglitazone selectively inhibits K(ATP) channels by acting on the K(IR) 6 subunit
Abstract
Background and purpose: Rosiglitazone is an anti-diabetic drug acting as an insulin sensitizer. We recently found that rosiglitazone also inhibits the vascular isoform of ATP-sensitive K(+) channels and compromises vasodilatory effects of β-adrenoceptor activation and pinacidil. As its potency for the channel inhibition is in the micromolar range, rosiglitazone may be used as an effective K(ATP) channel inhibitor for research and therapeutic purposes. Therefore, we performed experiments to determine whether other isoforms of K(ATP) channels are also sensitive to rosiglitazone and what their sensitivities are.
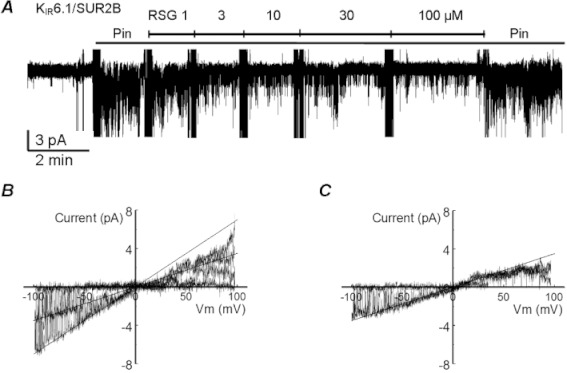
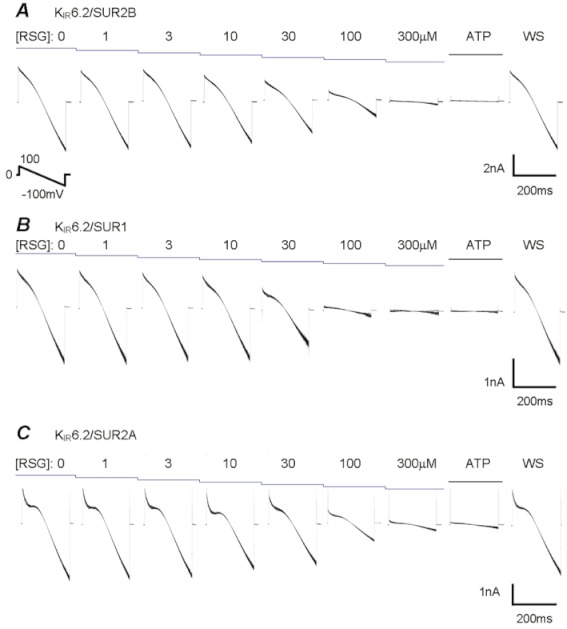
Experimental approach: K(IR) 6.1/SUR2B, K(IR) 6.2/SUR1, K(IR) 6.2/SUR2A, K(IR) 6.2/SUR2B and K(IR) 6.2ΔC36 channels were expressed in HEK293 cells and were studied using patch-clamp techniques.
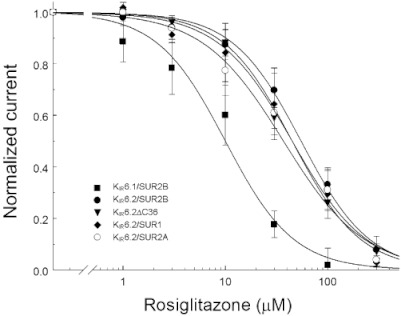
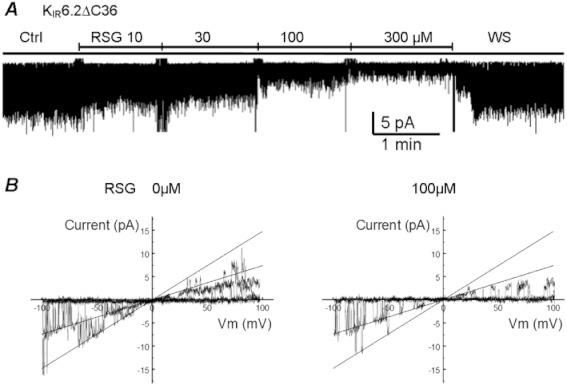
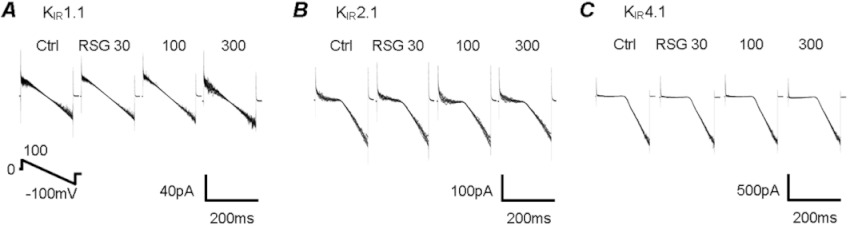
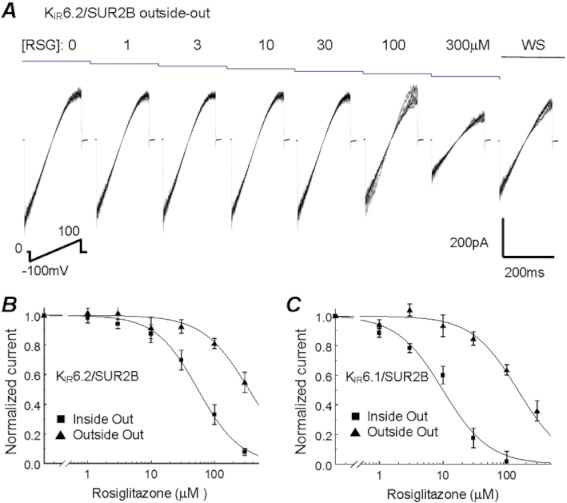
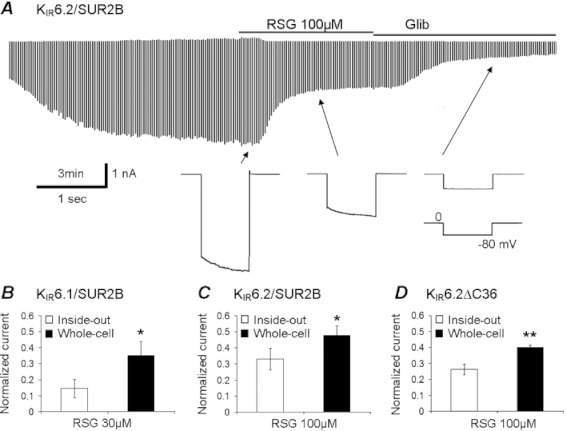
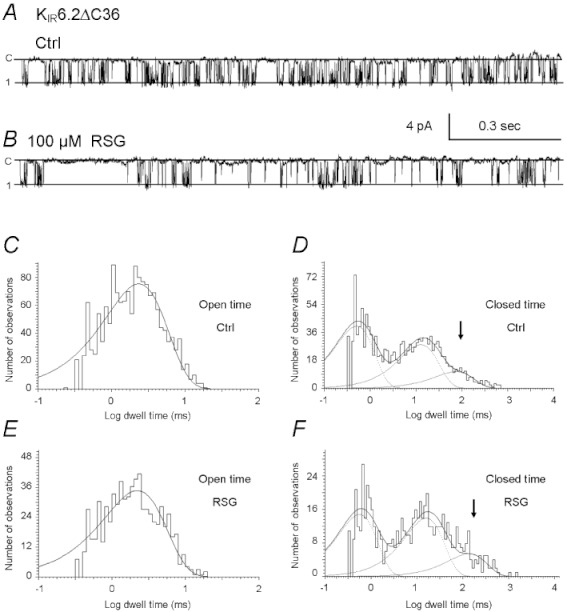
Key results: Rosiglitazone inhibited all isoforms of K(ATP) channels in excised patches and in the whole-cell configuration. Its IC(50) was 10 µmol·L(-1) for the K(IR) 6.1/SUR2B channel and ∼45 µmol·L(-1) for K(IR) 6.2/SURx channels. Rosiglitazone also inhibited K(IR) 6.2ΔC36 channels in the absence of the sulphonylurea receptor (SUR) subunit, with potency (IC(50) = 45 µmol·L(-1) ) almost identical to that for K(IR) 6.2/SURx channels. Single-channel kinetic analysis showed that the channel inhibition was mediated by augmentation of the long-lasting closures without affecting the channel open state and unitary conductance. In contrast, rosiglitazone had no effect on K(IR) 1.1, K(IR) 2.1 and K(IR) 4.1 channels, suggesting that the channel inhibitory effect is selective for K(IR) 6.x channels.
Conclusions and implications: These results suggest a novel K(ATP) channel inhibitor that acts on the pore-forming K(IR) 6.x subunit, affecting the channel gating.
© 2012 The Authors. British Journal of Pharmacology © 2012 The British Pharmacological Society.
Figures
Comment in
-
Selective block of K(ATP) channels: why the anti-diabetic sulphonylureas and rosiglitazone have more in common than we thought.Br J Pharmacol. 2012 Sep;167(1):23-5. doi: 10.1111/j.1476-5381.2012.01990.x. Br J Pharmacol. 2012. PMID: 22506686 Free PMC article.
References
-
- Abe M, Takiguchi Y, Ichimaru S, Kaji S, Tsuchiya K, Wada K. Different effect of acute treatment with rosiglitazone on rat myocardial ischemia/reperfusion injury by administration method. Eur J Pharmacol. 2008;589:215–219. - PubMed
-
- Ashcroft FM. From molecule to malady. Nature. 2006;440:440–447. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
