

Retinoic acid upregulates preadipocyte genes to block adipogenesis and suppress diet-induced obesity
- PMID: 22396202
- PMCID: PMC3331760
- DOI: 10.2337/db11-1620
Retinoic acid upregulates preadipocyte genes to block adipogenesis and suppress diet-induced obesity
Abstract
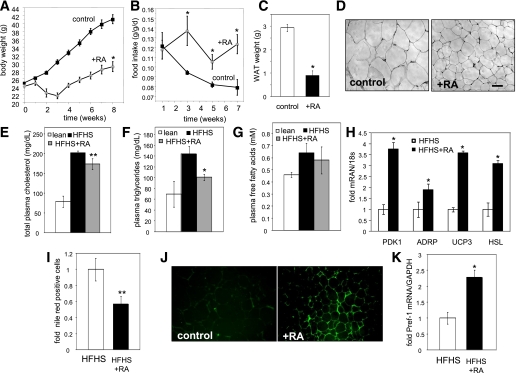
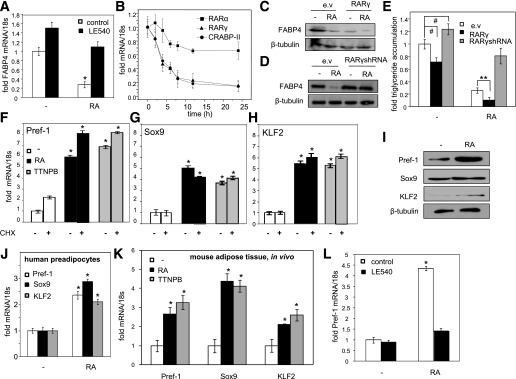
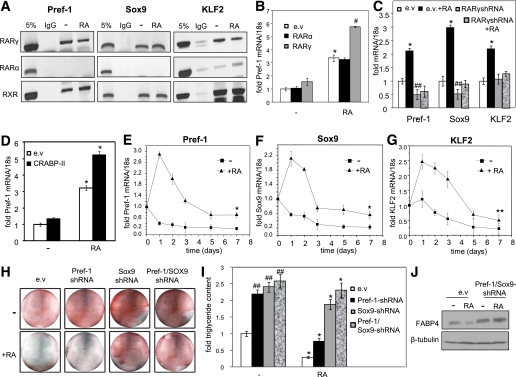
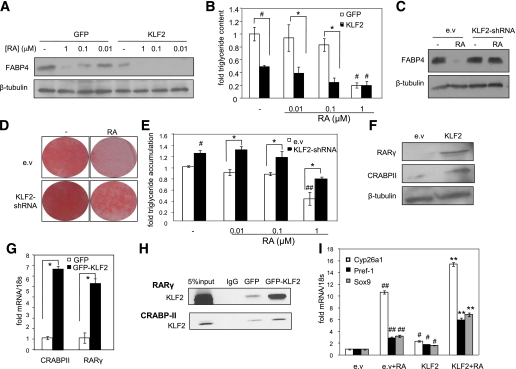
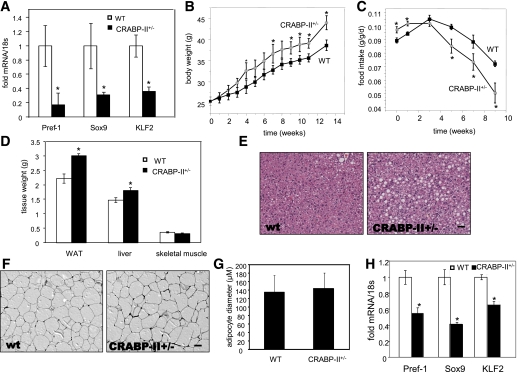
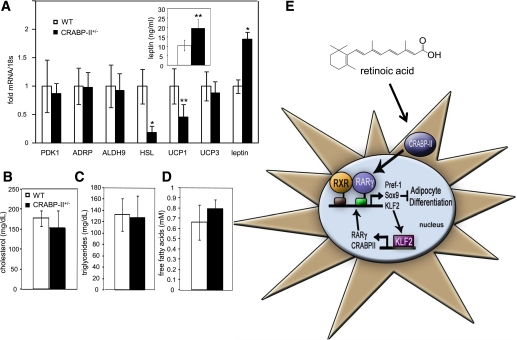
Retinoic acid (RA) protects mice from diet-induced obesity. The activity is mediated in part through activation of the nuclear receptors RA receptors (RARs) and peroxisome proliferator-activated receptor β/δ and their associated binding proteins cellular RA binding protein type II (CRABP-II) and fatty acid binding protein type 5 in adipocytes and skeletal muscle, leading to enhanced lipid oxidation and energy dissipation. It was also reported that RA inhibits differentiation of cultured preadipocytes. However, whether the hormone suppresses adipogenesis in vivo and how the activity is propagated remained unknown. In this study, we show that RA inhibits adipocyte differentiation by activating the CRABP-II/RARγ path in preadipose cells, thereby upregulating the expression of the adipogenesis inhibitors Pref-1, Sox9, and Kruppel-like factor 2 (KLF2). In turn, KLF2 induces the expression of CRABP-II and RARγ, further potentiating inhibition of adipocyte differentiation by RA. The data also indicate that RA suppresses adipogenesis in vivo and that the activity significantly contributes to the ability of the hormone to counteract diet-induced obesity.
Figures
References
-
- Green H, Kehinde O. An established preadipose cell line and its differentiation in culture. II. Factors affecting the adipose conversion. Cell 1975;5:19–27 - PubMed
-
- Green H, Meuth M. An established pre-adipose cell line and its differentiation in culture. Cell 1974;3:127–133 - PubMed
-
- Tontonoz P, Hu E, Graves RA, Budavari AI, Spiegelman BM. mPPAR gamma 2: tissue-specific regulator of an adipocyte enhancer. Genes Dev 1994;8:1224–1234 - PubMed
-
- Shao D, Lazar MA. Peroxisome proliferator activated receptor gamma, CCAAT/enhancer-binding protein alpha, and cell cycle status regulate the commitment to adipocyte differentiation. J Biol Chem 1997;272:21473–21478 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
