

Review
doi: 10.1523/JNEUROSCI.5372-11.2012.
Sex differences in the brain: the not so inconvenient truth
Affiliations
- PMID: 22396398
- PMCID: PMC3295598
- DOI: 10.1523/JNEUROSCI.5372-11.2012
Item in Clipboard
Review
Sex differences in the brain: the not so inconvenient truth
J Neurosci.
.
No abstract available
Figures
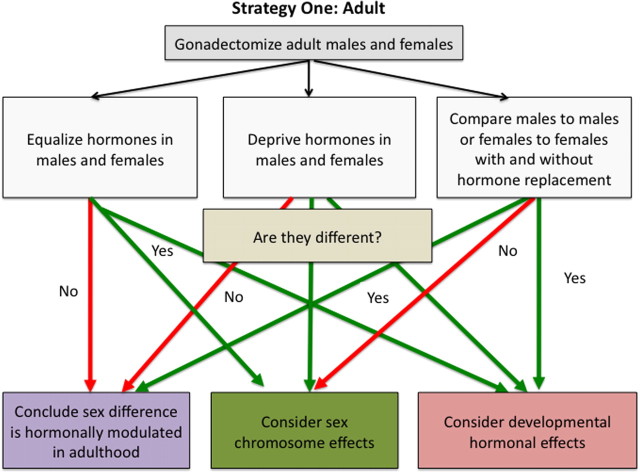
Strategy for discerning the basis of a sex difference starting with adult difference in the activational actions of testicular versus ovarian hormones. Because sex differences caused by gonadal hormones are the majority and are easier to study than those caused by sex chromosome effects, a simple way to start is to remove the gonads of males and females as adults and then perform various comparisons. If the sex difference persists when all gonadal hormones are removed, the source of the sex difference may be organizational effects of steroids during development or differences in sex chromosome complement. A second useful approach is to provide exogenous hormones to gonadectomized animals. If evidence suggests a sex difference is male biased, then starting by mimicking the hormonal status of males in both sexes (i.e., both receive testosterone) is probably the best approach. Conversely, if there is reason to believe a sex difference is female biased, then begin by providing both sexes a female typical hormonal profile (i.e., both receive estradiol or estradiol plus progesterone). If equating hormone levels abolishes the sex difference, the sex difference is due to adult differences in gonadal steroid levels. If, however, the sex difference persists, one should again consider the effects of developmental hormone exposure or sex chromosome complement.
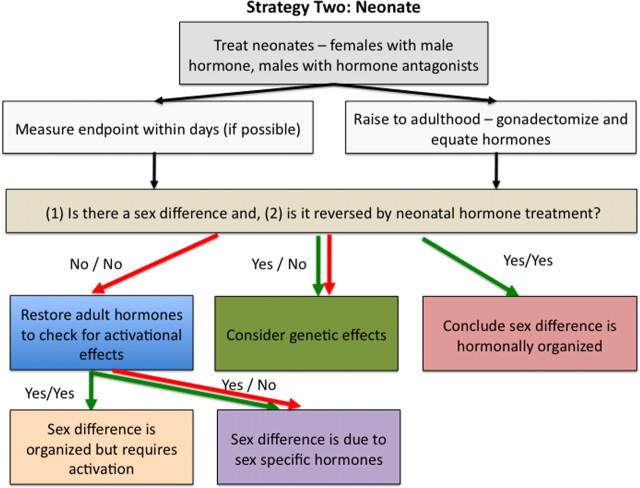
Strategy for discerning the basis of a sex difference starting with the neonate. Because sex differences in the action of gonadal hormones cause permanent sex differences in brain circuits, for some endpoints it is acceptable to test immediately for such fetal or neonatal effects. Following well established protocols available in the work of Becker et al. (2005), neonatal rats and mice can be readily treated with steroid hormones. Comparing males, females, males treated with blockers of steroid hormone action, and females administered masculinizing doses of steroid hormones tests whether a sex difference in hormone action during development causes the sex difference. If a particular endpoint cannot be measured in the neonate, animals are raised to adulthood. Because neonatal treatment can alter adult levels of gonadal hormones by altering the hypothalamic-pituitary-gonadal axis, any effect of neonatal treatment could be interpreted as a change in brain circuits or an altered hormonal profile if the gonads remain. A common method to distinguish between these possibilities is to remove group differences in hormone levels in adulthood by gonadectomy and, if possible, to compare males and females on the relevant behavioral or neural variable. If the sex difference persists, this suggests either a purely organizational effect with no role for hormonal activation, or a genetic basis for the sex difference. In many cases, however, the endpoint under study will not be evident in conditions in which hormones are completely lacking. In these instances the appropriate approach is treating males and females with similar hormones and then observing their response. If a sex difference is observed either developmentally or in adulthood and the sex difference is reversed by hormone treatment of the fetus or neonate, one can conclude the sex difference was hormonally “organized.” If the sex difference remains, direct sex chromosome effects may contribute to the sex difference. In some instances the treatment of neonatal males with hormone blockers has no effect. This can be due to the masculinizing effects of prenatal testicular secretions, and therefore cannot be disrupted by postnatal blockade of hormone effects. This possibility can be further explored by prenatal treatments or other tools (e.g., gene knock-out mice).
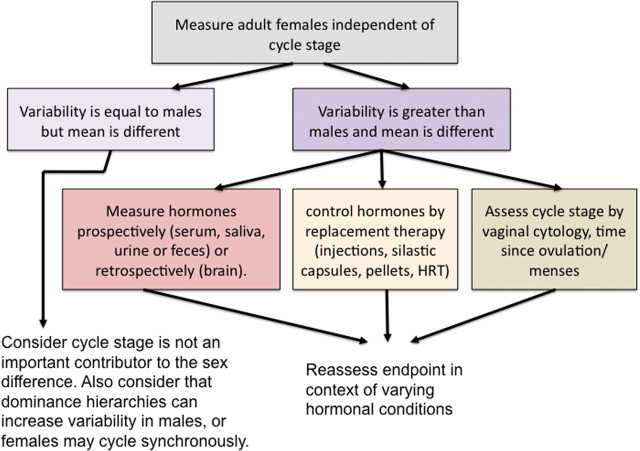
Impact of female cyclical reproduction on sex differences. Many endpoints with robust sex differences are not modulated by reproductive cycle stage, and a strategy is to assume this is the case until proven otherwise. A simple comparison of gonadally intact males and females will reveal whether there is greater variability in the females compared with the males. If so, this suggests that reproductive cycle stage might be important. There are various approaches that can be used to either determine the status of naturally cycling females or to provide them with a standard regimen of exogenous hormones. If the variability in females does not differ from that in males, this does not necessarily mean there is no role for the estrous cycle. Variability in males may be increased by unanticipated factors such as dominance status during group housing. Conversely, variability in females may be reduced because of either suppression or synchronization of reproductive cycles within group-housed females. Because assessing the stage of the cycle can be laborious, a simpler strategy may be to first determine which hormone(s) when given exogenously alters the endpoint, and then decide whether further assessing estrous cycle stage is worthwhile.
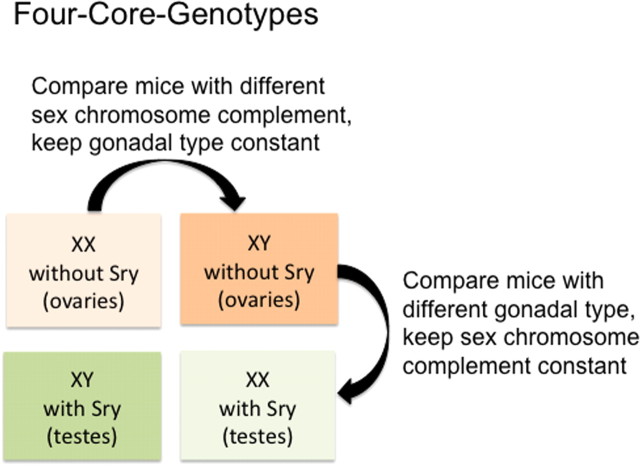
One strategy for detecting a “direct sex chromosome effect” on a sex difference. The importance of genetic effects to sex differences in the brain is becoming increasingly evident, and there is now a strong strategy with which to study them. The advent of the Four-Core-Genotypes mouse model provides an opportunity to compare mice of the same sex chromosome complement (XX versus XX) but of the opposite gonadal phenotype (ovaries versus testes). Thus, if an endpoint varies by gonadal phenotype, it is considered hormonal in origin, and if it varies by chromosome complement, it is considered genetic in origin. There is also the potential for interactive effects between gonadal phenotype and chromosome complement.
References
-
- Abel KM, Drake R, Goldstein JM. Sex differences in schizophrenia. Int Rev Psychiatry. 2010;22:417–428. - PubMed
-
- Amateau SK, McCarthy MM. Induction of PGE(2) by estradiol mediates developmental masculinization of sex behavior. Nat Neurosci. 2004;7:643–650. - PubMed
-
- Arnold AP, Wade J, Grisham W, Jacobs EC, Campagnoni AT. Sexual differentiation of the brain in songbirds. Dev Neurosci. 1996;18:124–136. - PubMed
-
- Arnold AP, Burgoyne PS. Are XX and XY brain cells intrinsically different? Trends Endocrinol Metab. 2004;15:6–11. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources