

Meiotic competent human germ cell-like cells derived from human embryonic stem cells induced by BMP4/WNT3A signaling and OCT4/EpCAM (epithelial cell adhesion molecule) selection
- PMID: 22396540
- PMCID: PMC3340282
- DOI: 10.1074/jbc.M111.338434
Meiotic competent human germ cell-like cells derived from human embryonic stem cells induced by BMP4/WNT3A signaling and OCT4/EpCAM (epithelial cell adhesion molecule) selection
Abstract
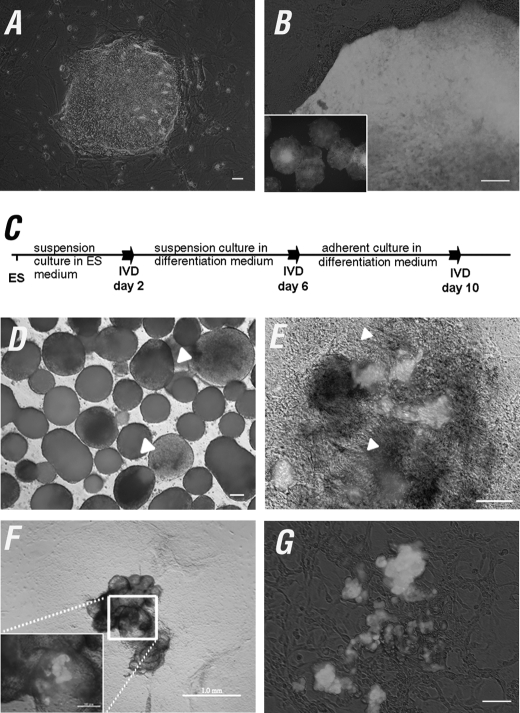
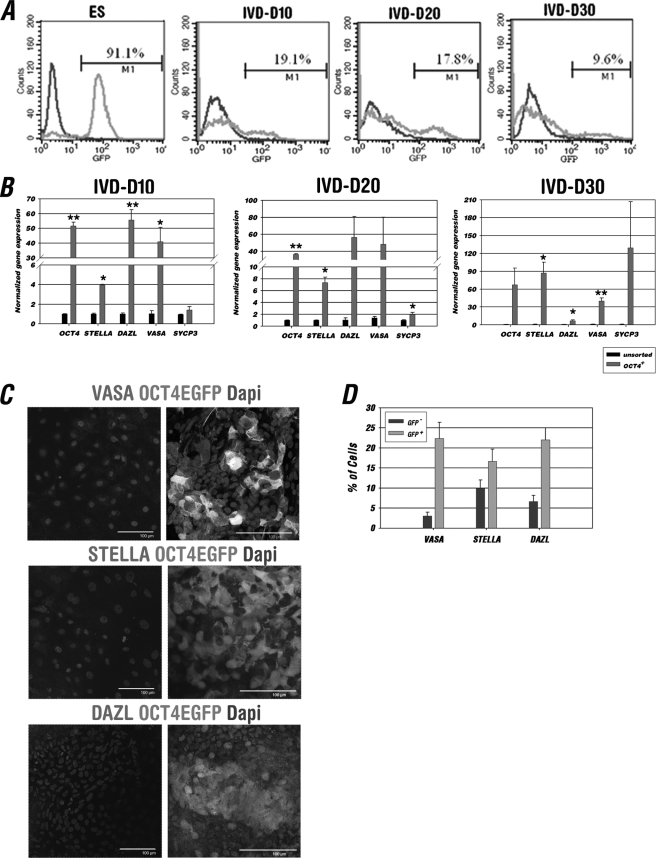
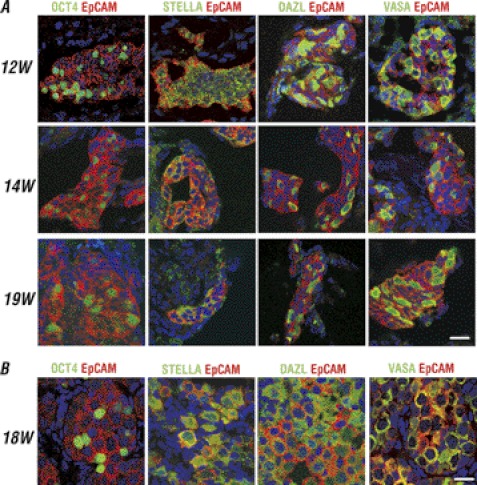
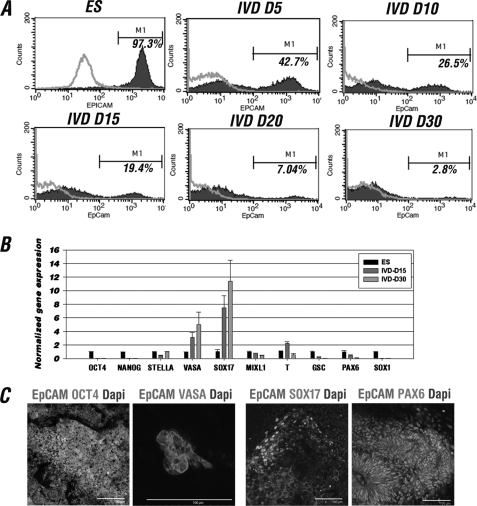
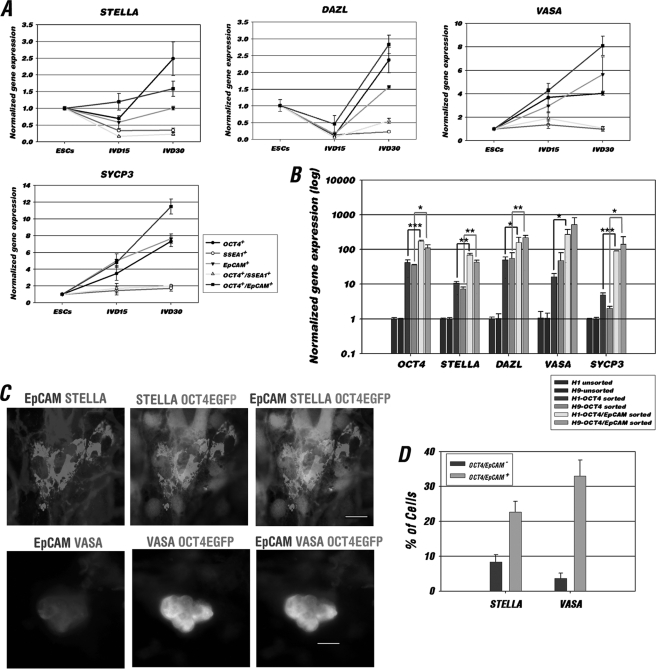
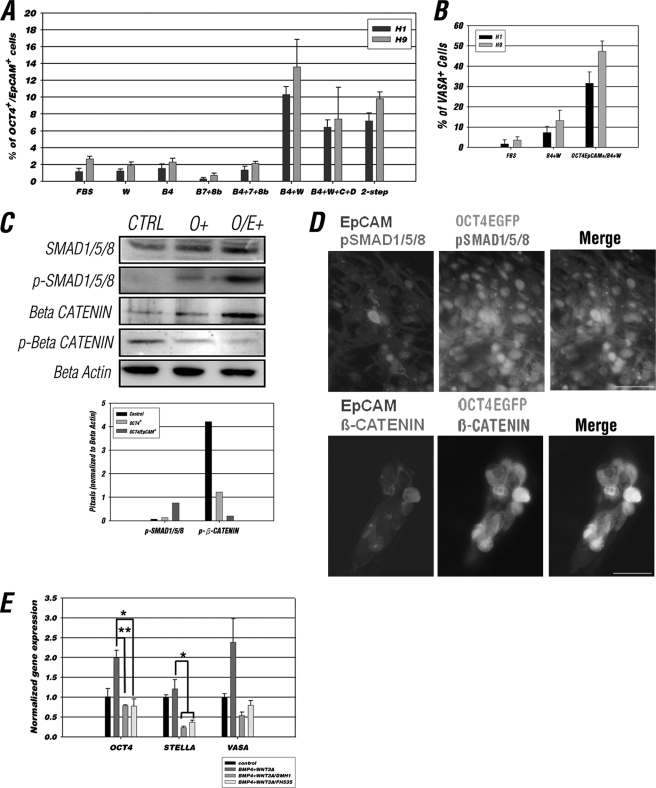
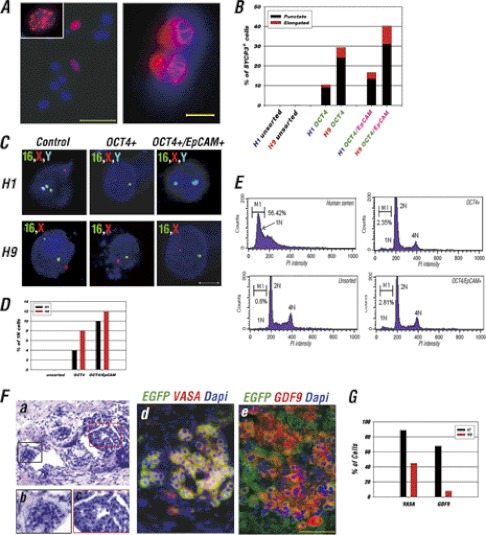
The establishment of an effective germ cell selection/enrichment platform from in vitro differentiating human embryonic stem cells (hESCs) is crucial for studying the molecular and signaling processes governing human germ cell specification and development. In this study, we developed a germ cell-enriching system that enables us to identify signaling factors involved in germ cell-fate induction from differentiating hESCs in vitro. First, we demonstrated that selection through an OCT4-EGFP reporter system can successfully increase the percentage of meiotic-competent, germ cell-like cells from spontaneously differentiating hESCs. Furthermore, we showed that the pluripotency associated surface marker, epithelial cell adhesion molecule (EpCAM), is also expressed in human fetal gonads and can be used as an effective selection marker for germ cell enrichment from differentiating hESCs. Combining OCT4 and EpCAM selection can further enrich the meiotic-competent germ cell-like cell population. Also, with the percentage of OCT4(+)/EpCAM(+) cells as readout, we demonstrated the synergistic effect of BMP4/pSMAD1/5/8 and WNT3A/β-CATENIN in promoting hESCs toward the germline fate. Combining BMP4/WNT3A induction and OCT4/EpCAM selection can significantly increase the putative germ cell population with meiotic competency. Co-transplantation of these cells with dissociated mouse neonatal ovary cells into SCID mice resulted in a homogenous germ cell cluster formation in vivo. The stepwise platform established in this study provides a useful tool to elucidate the molecular mechanisms of human germ cell development, which has implications not only for human fertility research but regenerative medicine in general.
Figures
References
-
- Hübner K., Fuhrmann G., Christenson L. K., Kehler J., Reinbold R., De La Fuente R., Wood J., Strauss J. F., 3rd, Boiani M., Schöler H. R. (2003) Derivation of oocytes from mouse embryonic stem cells. Science 300, 1251–1256 - PubMed
-
- Nayernia K., Nolte J., Michelmann H. W., Lee J. H., Rathsack K., Drusenheimer N., Dev A., Wulf G., Ehrmann I. E., Elliott D. J., Okpanyi V., Zechner U., Haaf T., Meinhardt A., Engel W. (2006) In vitro differentiated embryonic stem cells give rise to male gametes that can generate offspring mice. Dev. Cell 11, 125–132 - PubMed
-
- Geijsen N., Horoschak M., Kim K., Gribnau J., Eggan K., Daley G. Q. (2004) Derivation of embryonic germ cells and male gametes from embryonic stem cells. Nature 427, 148–154 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
