

Redundant Notch1 and Notch2 signaling is necessary for IFNγ secretion by T helper 1 cells during infection with Leishmania major
- PMID: 22396647
- PMCID: PMC3291656
- DOI: 10.1371/journal.ppat.1002560
Redundant Notch1 and Notch2 signaling is necessary for IFNγ secretion by T helper 1 cells during infection with Leishmania major
Abstract
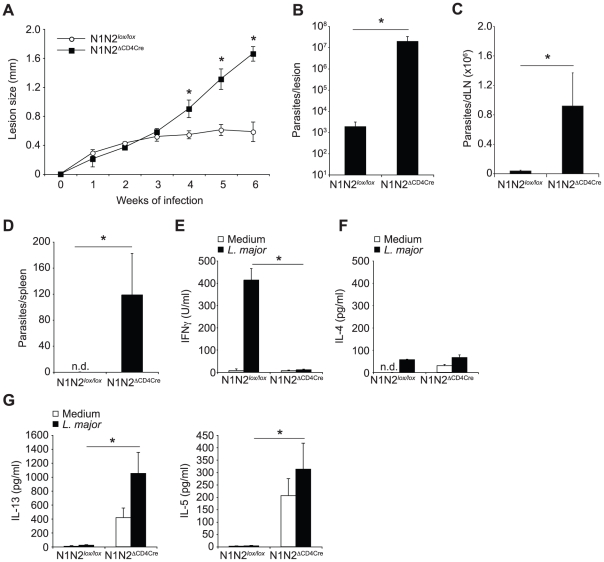
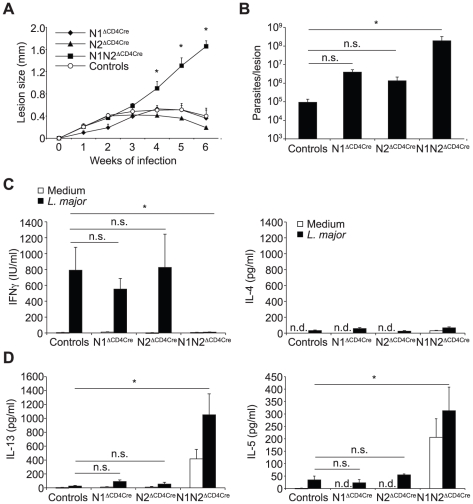
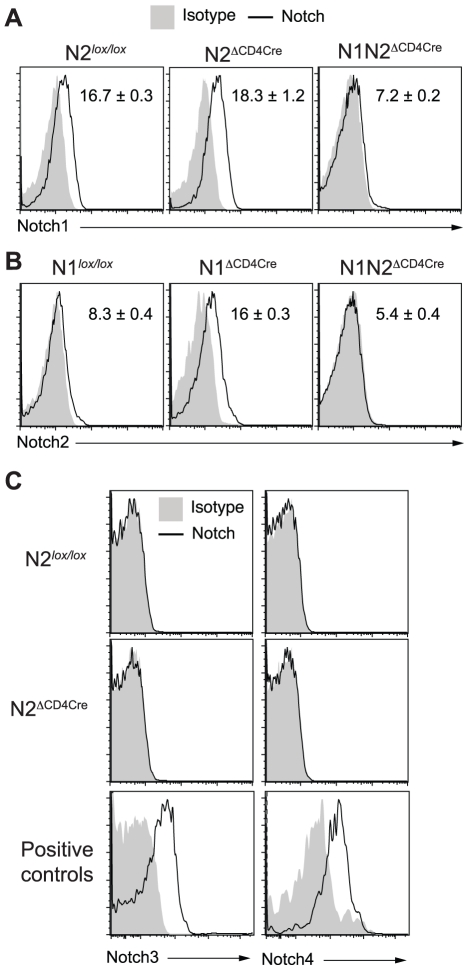
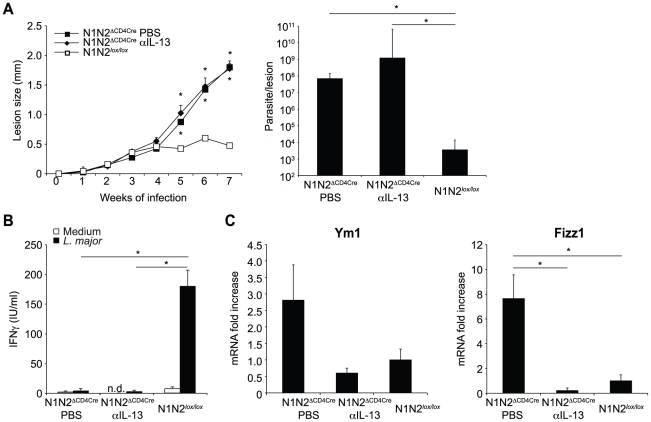
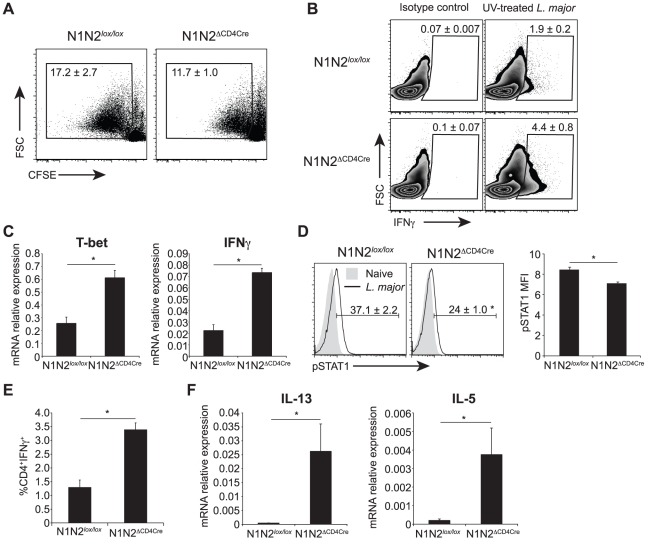
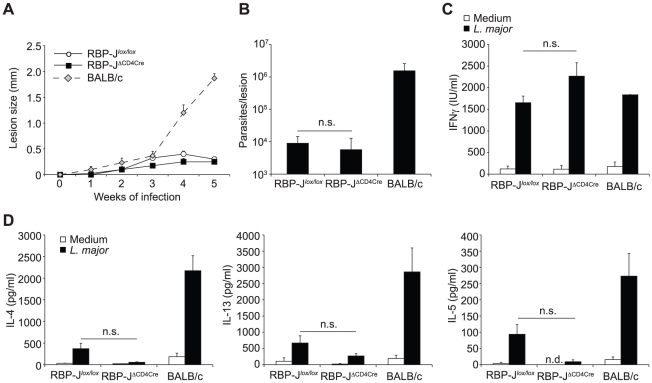
The protective immune response to intracellular parasites involves in most cases the differentiation of IFNγ-secreting CD4(+) T helper (Th) 1 cells. Notch receptors regulate cell differentiation during development but their implication in the polarization of peripheral CD4(+) T helper 1 cells is not well understood. Of the four Notch receptors, only Notch1 (N1) and Notch2 (N2) are expressed on activated CD4(+) T cells. To investigate the role of Notch in Th1 cell differentiation following parasite infection, mice with T cell-specific gene ablation of N1, N2 or both (N1N2(ΔCD4Cre)) were infected with the protozoan parasite Leishmania major. N1N2(ΔCD4Cre) mice, on the C57BL/6 L. major-resistant genetic background, developed unhealing lesions and uncontrolled parasitemia. Susceptibility correlated with impaired secretion of IFNγ by draining lymph node CD4(+) T cells and increased secretion of the IL-5 and IL-13 Th2 cytokines. Mice with single inactivation of N1 or N2 in their T cells were resistant to infection and developed a protective Th1 immune response, showing that CD4(+) T cell expression of N1 or N2 is redundant in driving Th1 differentiation. Furthermore, we show that Notch signaling is required for the secretion of IFNγ by Th1 cells. This effect is independent of CSL/RBP-Jκ, the major effector of Notch receptors, since L. major-infected mice with a RBP-Jκ deletion in their T cells were able to develop IFNγ-secreting Th1 cells, kill parasites and heal their lesions. Collectively, we demonstrate here a crucial role for RBP-Jκ-independent Notch signaling in the differentiation of a functional Th1 immune response following L. major infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Radtke F, Fasnacht N, Macdonald HR. Notch signaling in the immune system. Immunity. 2010;32:14–27. - PubMed
-
- Sandy AR, Maillard I. Notch signaling in the hematopoietic system. Expert Opin Biol Ther. 2009;9:1383–1398. - PubMed
-
- Osipo C, Golde TE, Osborne BA, Miele LA. Off the beaten pathway: the complex cross talk between Notch and NF-kappaB. Lab Invest. 2008;88:11–17. - PubMed
-
- Amsen D, Antov A, Flavell RA. The different faces of Notch in T-helper-cell differentiation. Nat Rev Immunol. 2009;9:116–124. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
