

Insulin/IGF-1 and hypoxia signaling act in concert to regulate iron homeostasis in Caenorhabditis elegans
- PMID: 22396654
- PMCID: PMC3291539
- DOI: 10.1371/journal.pgen.1002498
Insulin/IGF-1 and hypoxia signaling act in concert to regulate iron homeostasis in Caenorhabditis elegans
Abstract
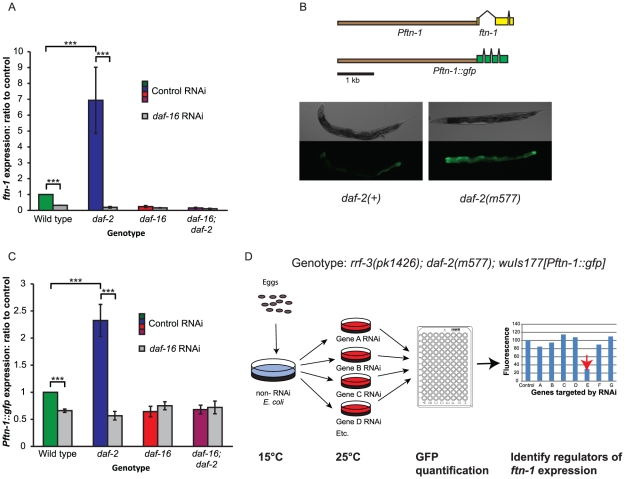
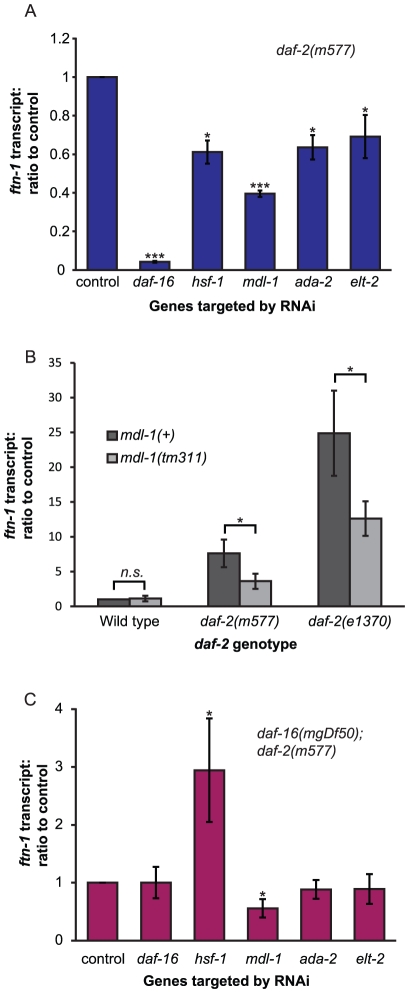
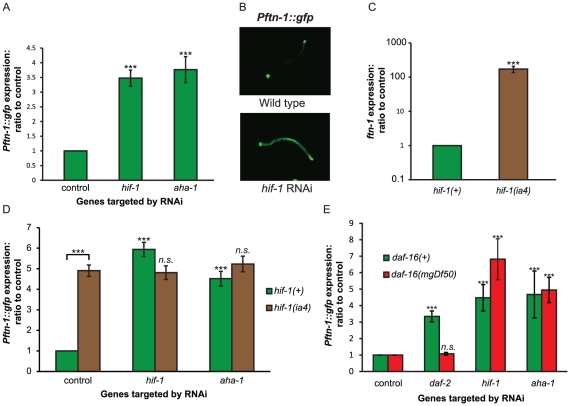
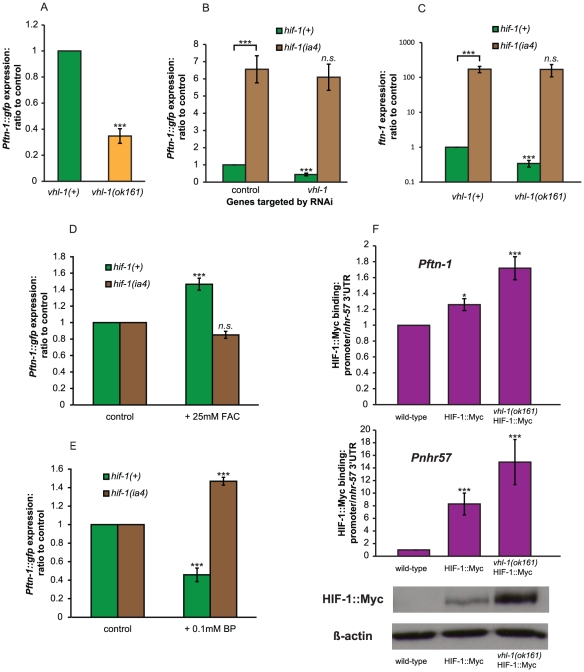
Iron plays an essential role in many biological processes, but also catalyzes the formation of reactive oxygen species (ROS), which can cause molecular damage. Iron homeostasis is therefore a critical determinant of fitness. In Caenorhabditis elegans, insulin/IGF-1 signaling (IIS) promotes growth and reproduction but limits stress resistance and lifespan through inactivation of the DAF-16/FoxO transcription factor (TF). We report that long-lived daf-2 insulin/IGF-1 receptor mutants show a daf-16-dependent increase in expression of ftn-1, which encodes the iron storage protein H-ferritin. To better understand the regulation of iron homeostasis, we performed a TF-limited genetic screen for factors influencing ftn-1 gene expression. The screen identified the heat-shock TF hsf-1, the MAD bHLH TF mdl-1, and the putative histone acetyl transferase ada-2 as activators of ftn-1 expression. It also revealed that the HIFα homolog hif-1 and its binding partner aha-1 (HIFβ) are potent repressors of ftn-1 expression. ftn-1 expression is induced by exposure to iron, and we found that hif-1 was required for this induction. In addition, we found that the prolyl hydroxylase EGL-9, which represses HIF-1 via the von Hippel-Lindau tumor suppressor VHL-1, can also act antagonistically to VHL-1 in regulating ftn-1. This suggests a novel mechanism for HIF target gene regulation by these evolutionarily conserved and clinically important hydroxylases. Our findings imply that the IIS and HIF pathways act together to regulate iron homeostasis in C. elegans. We suggest that IIS/DAF-16 regulation of ftn-1 modulates a trade-off between growth and stress resistance, as elevated iron availability supports growth but also increases ROS production.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
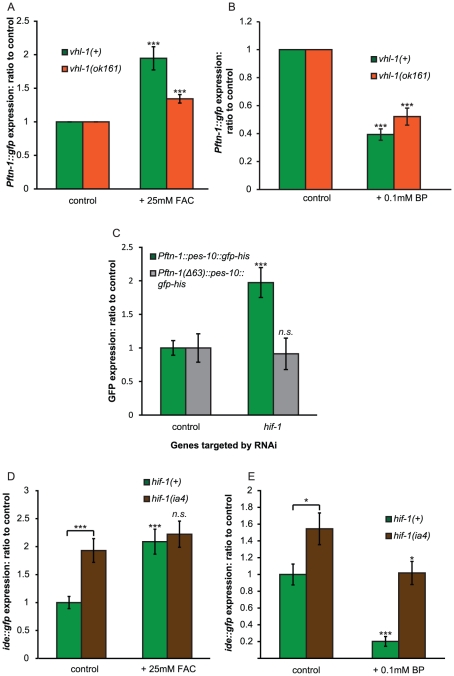
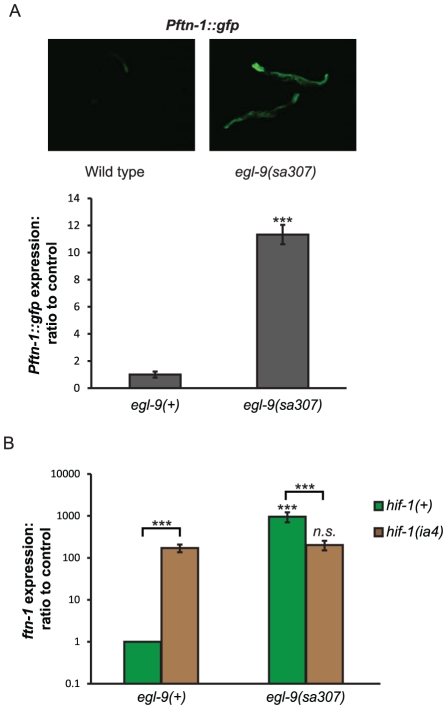
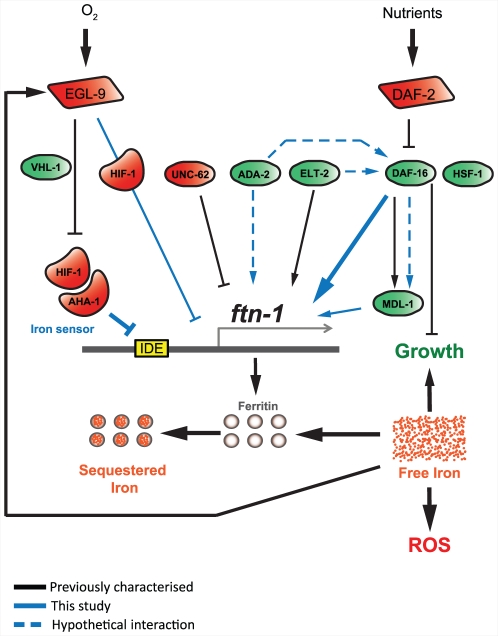
References
-
- Riddle DL, Albert PS. 1997. Genetic and Environmental Regulation of Dauer Larva Development.
-
- Cassada RC, Russell RL. The dauerlarva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Developmental Biology. 1975;46:326–342. - PubMed
-
- Lithgow GJ, Walker GA. Stress resistance as a determinate of C. elegans lifespan. Mechanisms of Ageing and Development. 2002;123:765–771. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
