

Identification of novel causative genes determining the complex trait of high ethanol tolerance in yeast using pooled-segregant whole-genome sequence analysis
- PMID: 22399573
- PMCID: PMC3337442
- DOI: 10.1101/gr.131698.111
Identification of novel causative genes determining the complex trait of high ethanol tolerance in yeast using pooled-segregant whole-genome sequence analysis
Abstract
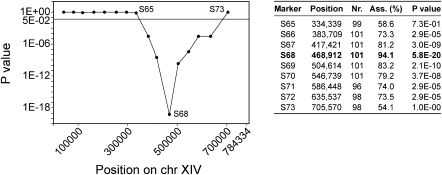
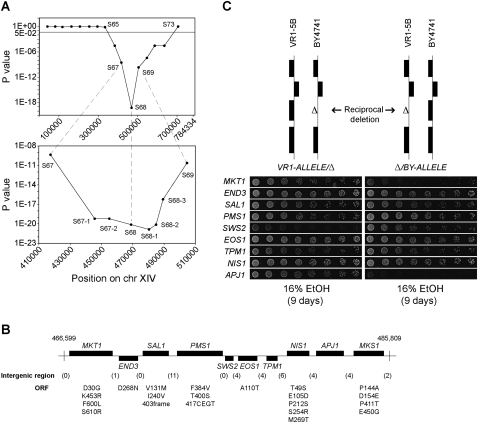
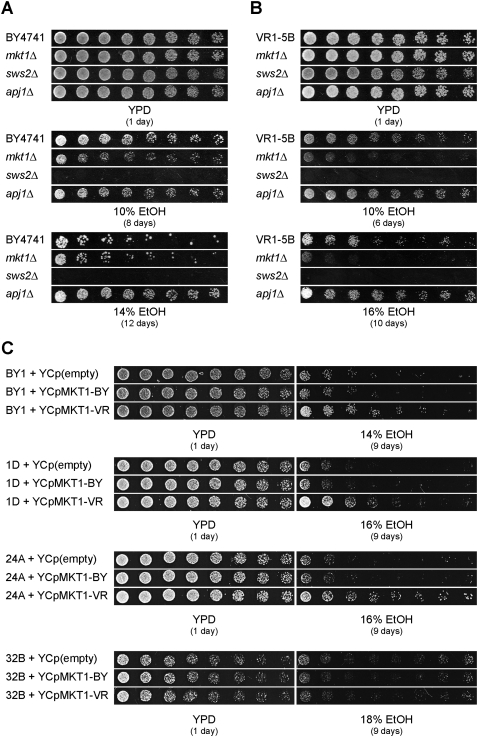
High ethanol tolerance is an exquisite characteristic of the yeast Saccharomyces cerevisiae, which enables this microorganism to dominate in natural and industrial fermentations. Up to now, ethanol tolerance has only been analyzed in laboratory yeast strains with moderate ethanol tolerance. The genetic basis of the much higher ethanol tolerance in natural and industrial yeast strains is unknown. We have applied pooled-segregant whole-genome sequence analysis to map all quantitative trait loci (QTL) determining high ethanol tolerance. We crossed a highly ethanol-tolerant segregant of a Brazilian bioethanol production strain with a laboratory strain with moderate ethanol tolerance. Out of 5974 segregants, we pooled 136 segregants tolerant to at least 16% ethanol and 31 segregants tolerant to at least 17%. Scoring of SNPs using whole-genome sequence analysis of DNA from the two pools and parents revealed three major loci and additional minor loci. The latter were more pronounced or only present in the 17% pool compared to the 16% pool. In the locus with the strongest linkage, we identified three closely located genes affecting ethanol tolerance: MKT1, SWS2, and APJ1, with SWS2 being a negative allele located in between two positive alleles. SWS2 and APJ1 probably contained significant polymorphisms only outside the ORF, and lower expression of APJ1 may be linked to higher ethanol tolerance. This work has identified the first causative genes involved in high ethanol tolerance of yeast. It also reveals the strong potential of pooled-segregant sequence analysis using relatively small numbers of selected segregants for identifying QTL on a genome-wide scale.
Figures


References
-
- Alexandre H, Ansanay-Galeote V, Dequin S, Blondin B 2001. Global gene expression during short-term ethanol stress in Saccharomyces cerevisiae. FEBS Lett 498: 98–103 - PubMed
-
- Brem RB, Yvert G, Clinton R, Kruglyak L 2002. Genetic dissection of transcriptional regulation in budding yeast. Science 296: 752–755 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases