

Claudin-17 forms tight junction channels with distinct anion selectivity
- PMID: 22402829
- PMCID: PMC11115135
- DOI: 10.1007/s00018-012-0949-x
Claudin-17 forms tight junction channels with distinct anion selectivity
Abstract
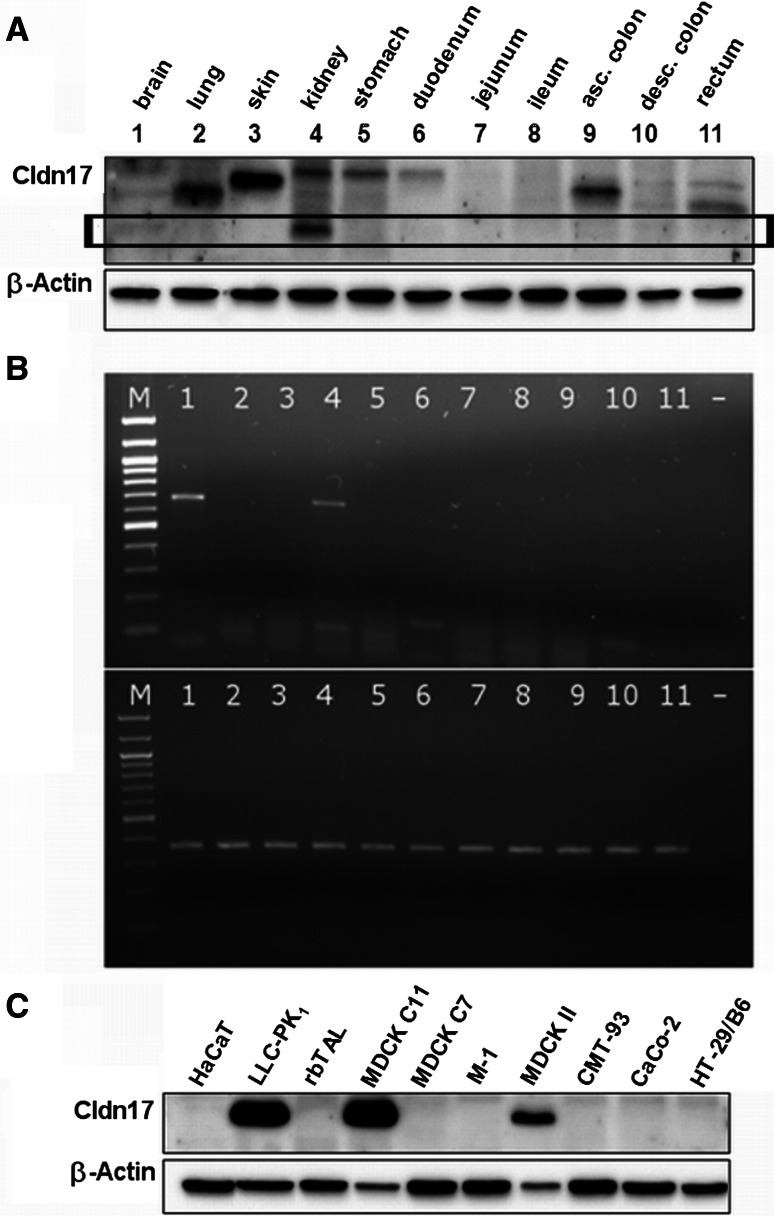
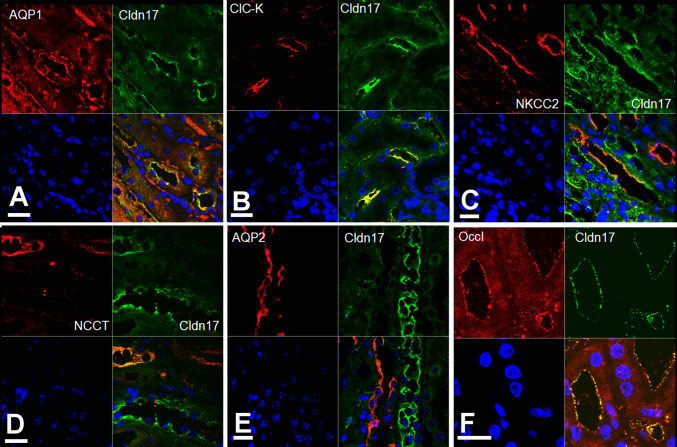
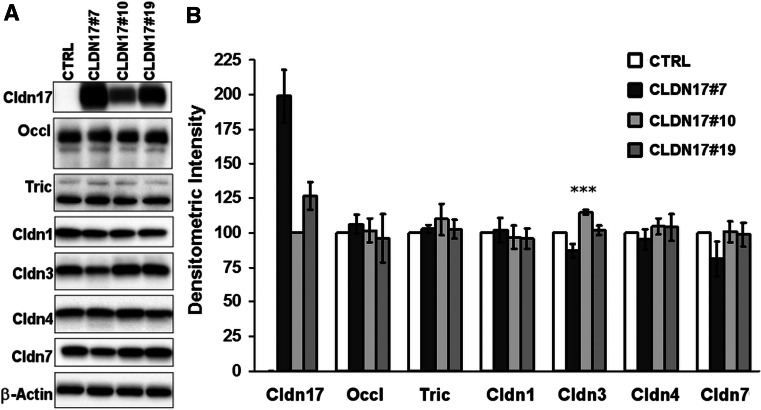
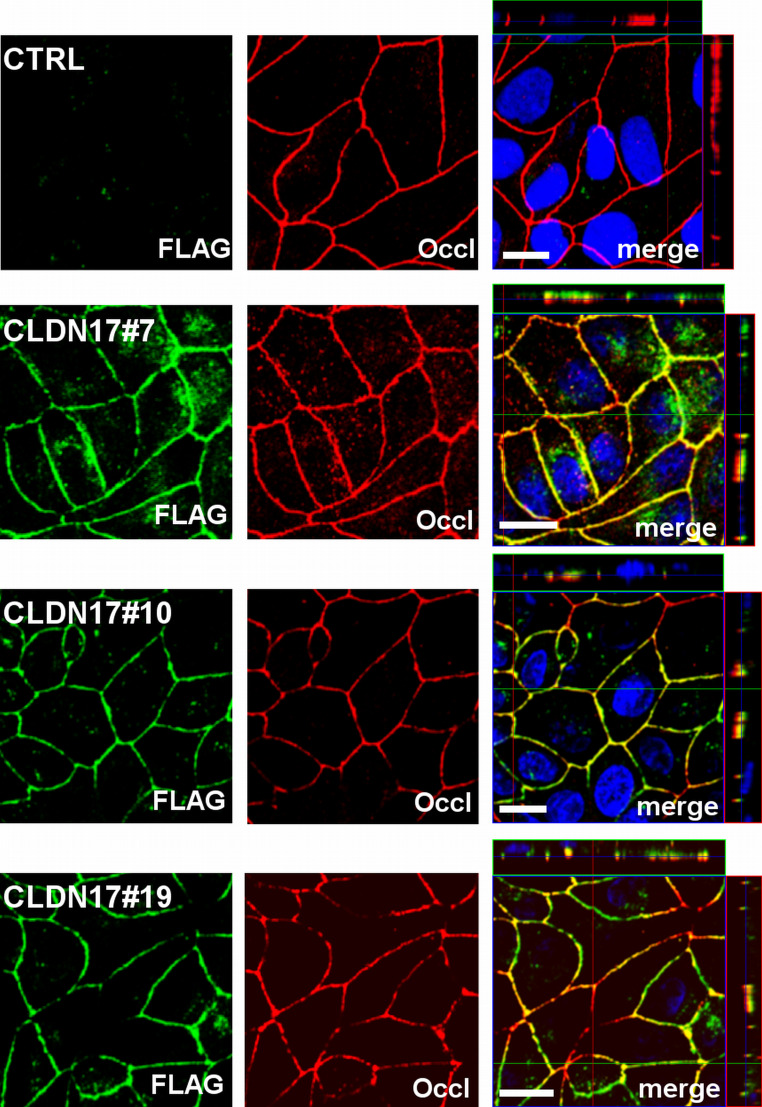
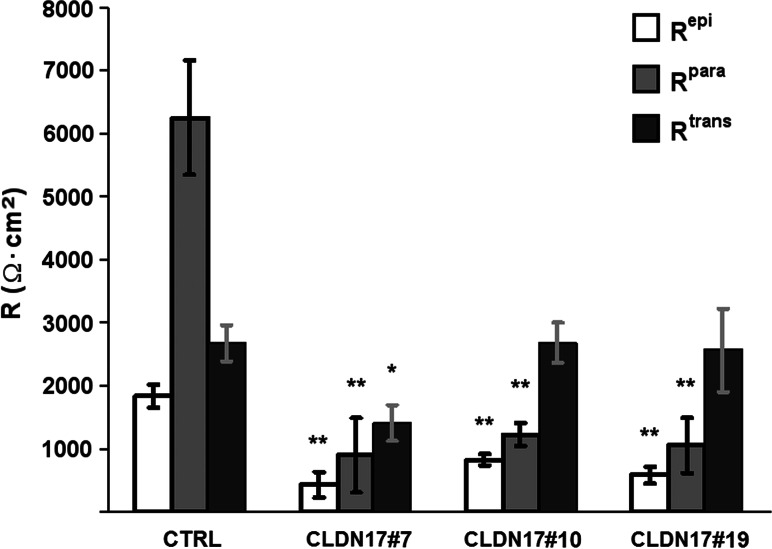
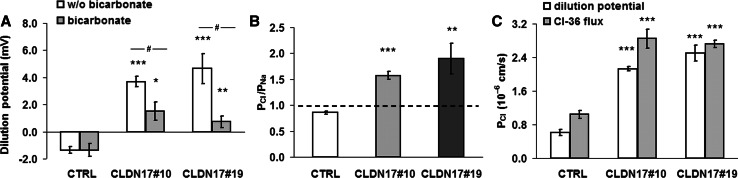
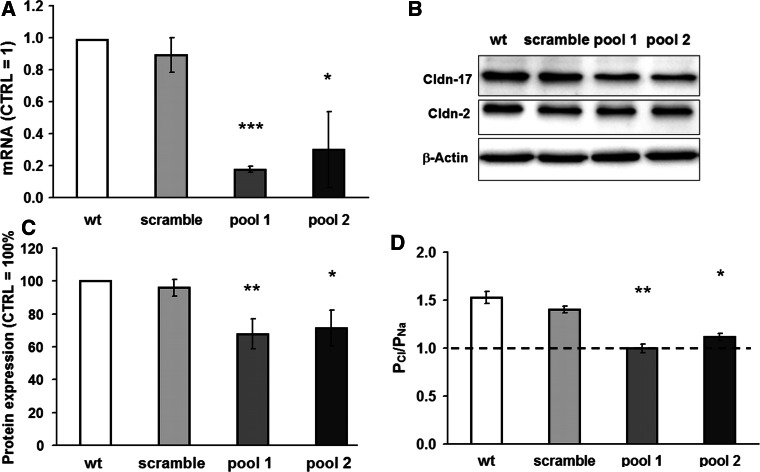
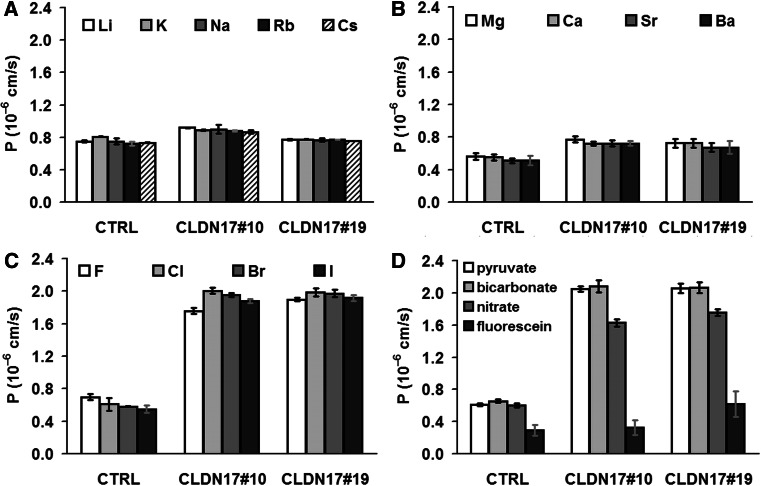
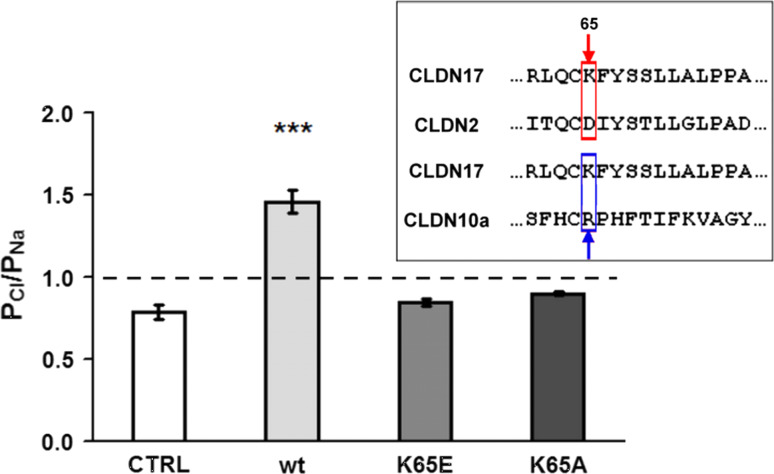
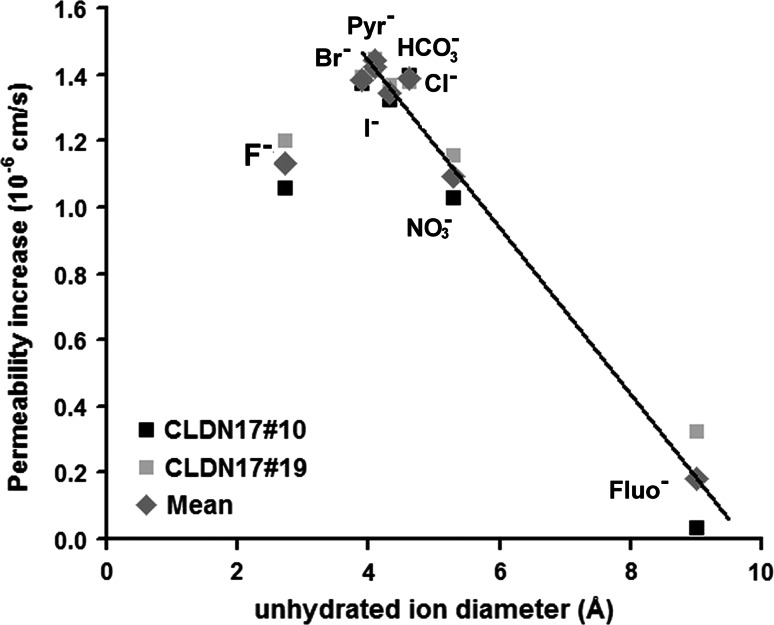
Barrier properties of tight junctions are determined by the claudin protein family. Many claudins seal this barrier, but others form paracellular channels. Among these, no claudins with general and clear-cut anion selectivity have yet been described, while for claudin-10a and claudin-4, only circumstantial or small anion selectivities have been shown. A claudin with unknown function and tissue distribution is claudin-17. We characterized claudin-17 by overexpression and knock-down in two renal cell lines. Overexpression in MDCK C7 cell layers caused a threefold increase in paracellular anion permeability and switched these cells from cation- to anion-selective. Knockdown in LLC-PK(1) cells indorsed the finding of claudin-17-based anion channels. Mutagenesis revealed that claudin-17 anion selectivity critically depends on a positive charge at position 65. Claudin-17 expression was found in two organs: marginal in brain but abundant in kidney, where expression was intense in proximal tubules and gradually decreased towards distal segments. As claudin-17 is predominantly expressed in proximal nephrons, which exhibit substantial, though molecularly not defined, paracellular chloride reabsorption, we suggest that claudin-17 has a unique physiological function in this process. In conclusion, claudin-17 forms channels within tight junctions with distinct anion preference.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures
Similar articles
-
Tight junctions of the proximal tubule and their channel proteins.Pflugers Arch. 2017 Aug;469(7-8):877-887. doi: 10.1007/s00424-017-2001-3. Epub 2017 Jun 9. Pflugers Arch. 2017. PMID: 28600680 Review.
-
Reversal of charge selectivity in cation or anion-selective epithelial lines by expression of different claudins.Am J Physiol Renal Physiol. 2003 Dec;285(6):F1078-84. doi: 10.1152/ajprenal.00116.2003. Epub 2003 Sep 16. Am J Physiol Renal Physiol. 2003. PMID: 13129853
-
Charge-selective claudin channels.Ann N Y Acad Sci. 2012 Jun;1257:20-8. doi: 10.1111/j.1749-6632.2012.06555.x. Ann N Y Acad Sci. 2012. PMID: 22671585
-
Claudin-10a Deficiency Shifts Proximal Tubular Cl- Permeability to Cation Selectivity via Claudin-2 Redistribution.J Am Soc Nephrol. 2022 Apr;33(4):699-717. doi: 10.1681/ASN.2021030286. Epub 2022 Jan 14. J Am Soc Nephrol. 2022. PMID: 35031570 Free PMC article.
-
Role of claudins in renal calcium handling.Nefrologia. 2015;35(4):347-52. doi: 10.1016/j.nefro.2015.06.011. Epub 2015 Jul 22. Nefrologia. 2015. PMID: 26306950 Review. English, Spanish.
Cited by
-
Bladder filling and voiding affect umbrella cell tight junction organization and function.Am J Physiol Renal Physiol. 2013 Oct 15;305(8):F1158-68. doi: 10.1152/ajprenal.00282.2013. Epub 2013 Jul 24. Am J Physiol Renal Physiol. 2013. PMID: 23884145 Free PMC article.
-
Epithelial integrity, junctional complexes, and biomarkers associated with intestinal functions.Tissue Barriers. 2022 Jul 3;10(3):1996830. doi: 10.1080/21688370.2021.1996830. Epub 2021 Oct 30. Tissue Barriers. 2022. PMID: 34719339 Free PMC article. Review.
-
The Basic Requirement of Tight Junction Proteins in Blood-Brain Barrier Function and Their Role in Pathologies.Int J Mol Sci. 2024 May 21;25(11):5601. doi: 10.3390/ijms25115601. Int J Mol Sci. 2024. PMID: 38891789 Free PMC article. Review.
-
Tight junctions of the proximal tubule and their channel proteins.Pflugers Arch. 2017 Aug;469(7-8):877-887. doi: 10.1007/s00424-017-2001-3. Epub 2017 Jun 9. Pflugers Arch. 2017. PMID: 28600680 Review.
-
Claudin-17 Deficiency in Mice Results in Kidney Injury Due to Electrolyte Imbalance and Oxidative Stress.Cells. 2022 May 29;11(11):1782. doi: 10.3390/cells11111782. Cells. 2022. PMID: 35681477 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
