

Control of protein signaling using a computationally designed GTPase/GEF orthogonal pair
- PMID: 22403064
- PMCID: PMC3325720
- DOI: 10.1073/pnas.1114487109
Control of protein signaling using a computationally designed GTPase/GEF orthogonal pair
Abstract
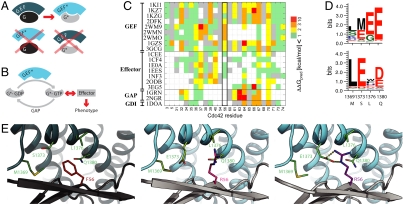
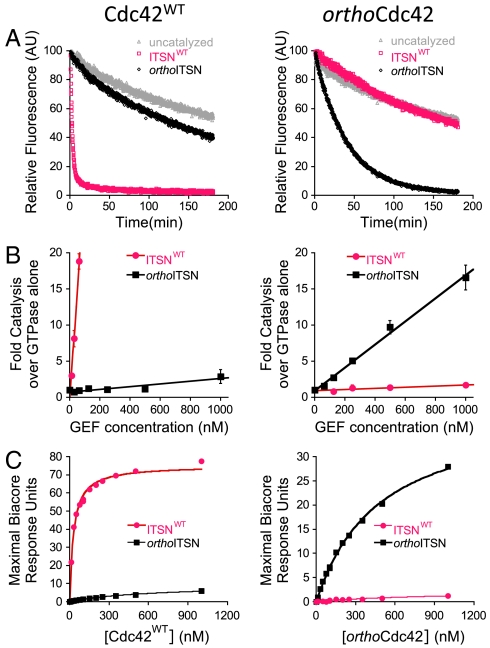
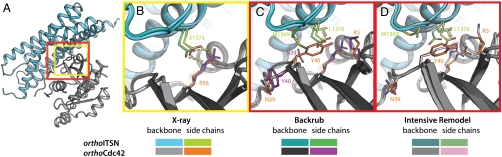
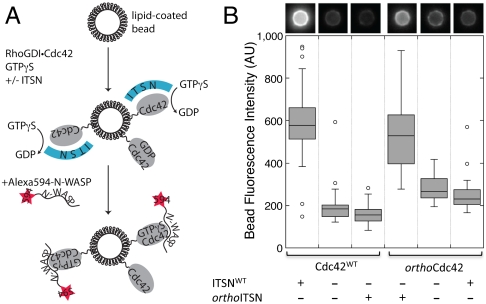
Signaling pathways depend on regulatory protein-protein interactions; controlling these interactions in cells has important applications for reengineering biological functions. As many regulatory proteins are modular, considerable progress in engineering signaling circuits has been made by recombining commonly occurring domains. Our ability to predictably engineer cellular functions, however, is constrained by complex crosstalk observed in naturally occurring domains. Here we demonstrate a strategy for improving and simplifying protein network engineering: using computational design to create orthogonal (non-crossreacting) protein-protein interfaces. We validated the design of the interface between a key signaling protein, the GTPase Cdc42, and its activator, Intersectin, biochemically and by solving the crystal structure of the engineered complex. The designed GTPase (orthoCdc42) is activated exclusively by its engineered cognate partner (orthoIntersectin), but maintains the ability to interface with other GTPase signaling circuit components in vitro. In mammalian cells, orthoCdc42 activity can be regulated by orthoIntersectin, but not wild-type Intersectin, showing that the designed interaction can trigger complex processes. Computational design of protein interfaces thus promises to provide specific components that facilitate the predictable engineering of cellular functions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
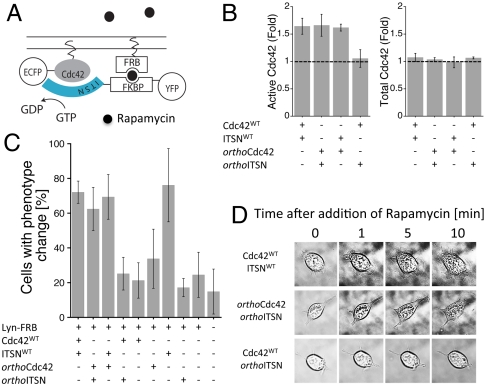
Comment in
-
Designing orthogonal signaling pathways: how to fit in with the surroundings.Proc Natl Acad Sci U S A. 2012 Apr 3;109(14):5140-1. doi: 10.1073/pnas.1202624109. Epub 2012 Mar 26. Proc Natl Acad Sci U S A. 2012. PMID: 22451928 Free PMC article. No abstract available.
References
-
- Elowitz MB, Leibler S. A synthetic oscillatory network of transcriptional regulators. Nature. 2000;403:335–338. - PubMed
-
- Gardner TS, Cantor CR, Collins JJ. Construction of a genetic toggle switch in Escherichia coli. Nature. 2000;403:339–342. - PubMed
-
- Sprinzak D, Elowitz MB. Reconstruction of genetic circuits. Nature. 2005;438:443–448. - PubMed
-
- Yeh BJ, Rutigliano RJ, Deb A, Bar-Sagi D, Lim WA. Rewiring cellular morphology pathways with synthetic guanine nucleotide exchange factors. Nature. 2007;447:596–600. - PubMed
-
- Bashor CJ, Helman NC, Yan S, Lim WA. Using engineered scaffold interactions to reshape MAP kinase pathway signaling dynamics. Science. 2008;319:1539–1543. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
