

Three acyltransferases and nitrogen-responsive regulator are implicated in nitrogen starvation-induced triacylglycerol accumulation in Chlamydomonas
- PMID: 22403401
- PMCID: PMC3346115
- DOI: 10.1074/jbc.M111.334052
Three acyltransferases and nitrogen-responsive regulator are implicated in nitrogen starvation-induced triacylglycerol accumulation in Chlamydomonas
Abstract
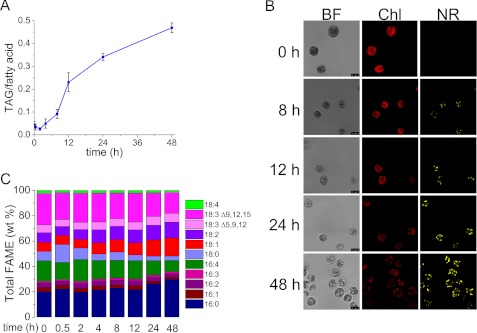
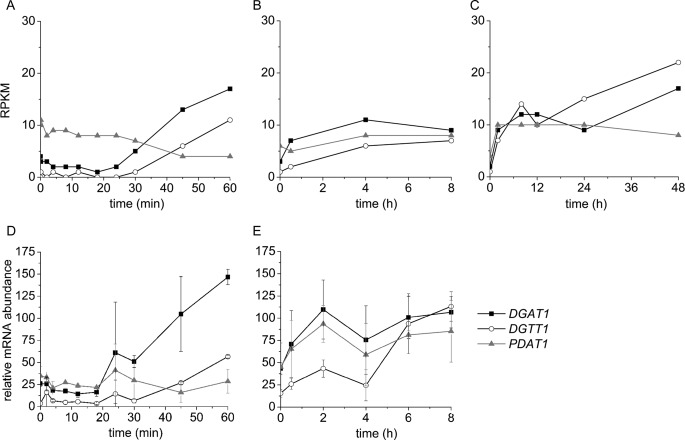
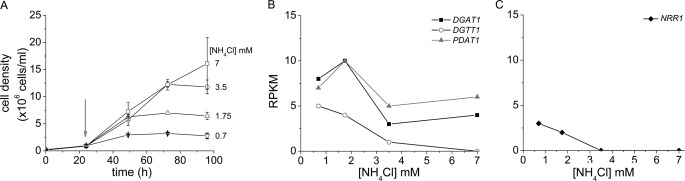
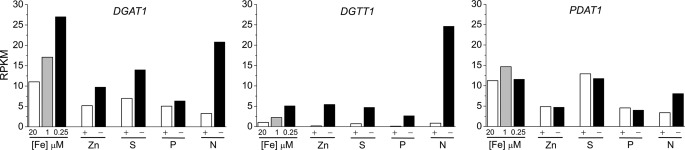
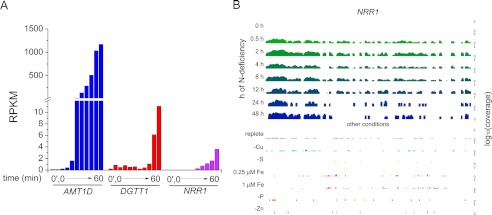
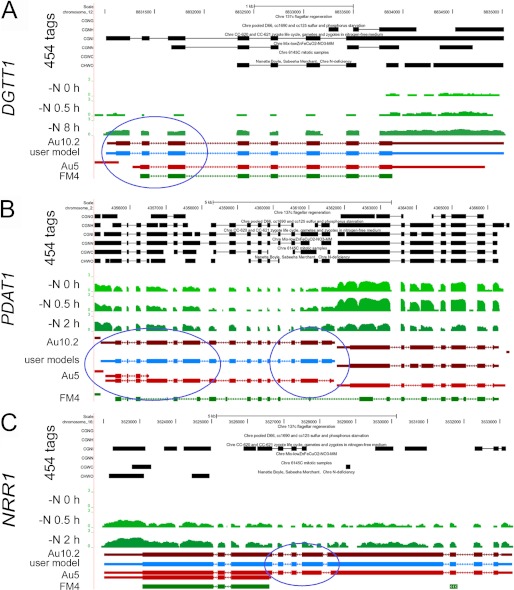
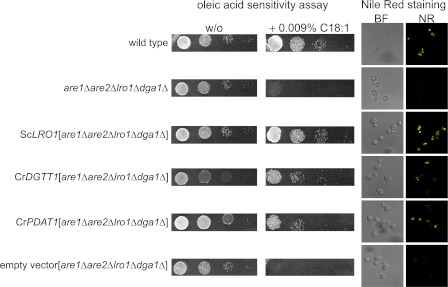
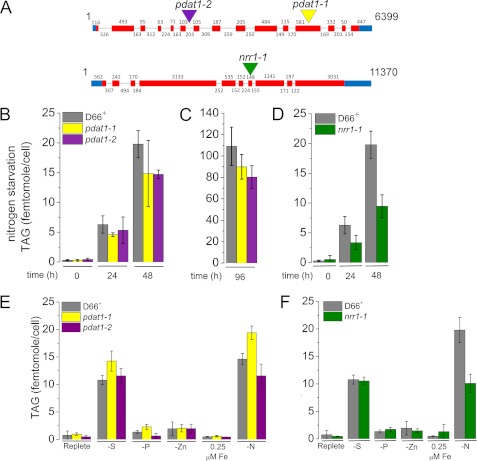
Algae have recently gained attention as a potential source for biodiesel; however, much is still unknown about the biological triggers that cause the production of triacylglycerols. We used RNA-Seq as a tool for discovering genes responsible for triacylglycerol (TAG) production in Chlamydomonas and for the regulatory components that activate the pathway. Three genes encoding acyltransferases, DGAT1, DGTT1, and PDAT1, are induced by nitrogen starvation and are likely to have a role in TAG accumulation based on their patterns of expression. DGAT1 and DGTT1 also show increased mRNA abundance in other TAG-accumulating conditions (minus sulfur, minus phosphorus, minus zinc, and minus iron). Insertional mutants, pdat1-1 and pdat1-2, accumulate 25% less TAG compared with the parent strain, CC-4425, which demonstrates the relevance of the trans-acylation pathway in Chlamydomonas. The biochemical functions of DGTT1 and PDAT1 were validated by rescue of oleic acid sensitivity and restoration of TAG accumulation in a yeast strain lacking all acyltransferase activity. Time course analyses suggest than a SQUAMOSA promoter-binding protein domain transcription factor, whose mRNA increases precede that of lipid biosynthesis genes like DGAT1, is a candidate regulator of the nitrogen deficiency responses. An insertional mutant, nrr1-1, accumulates only 50% of the TAG compared with the parental strain in nitrogen-starvation conditions and is unaffected by other nutrient stresses, suggesting the specificity of this regulator for nitrogen-deprivation conditions.
Figures
References
-
- Merchant S. S., Kropat J., Liu B., Shaw J., Warakanont J. (2012) TAG, You're it! Chlamydomonas as a reference organism for understanding algal triacylglycerol accumulation. Curr. Opin. Biotechnol. 23, 1–12 - PubMed
-
- Chisti Y. (2008) Biodiesel from microalgae beats bioethanol. Trends Biotechnol. 26, 126–131 - PubMed
-
- Chisti Y. (2007) Biodiesel from microalgae. Biotechnol. Adv. 25, 294–306 - PubMed
-
- Scott S. A., Davey M. P., Dennis J. S., Horst I., Howe C. J., Lea-Smith D. J., Smith A. G. (2010) Biodiesel from algae. Challenges and prospects. Curr. Opin. Biotechnol. 21, 277–286 - PubMed
-
- Schenk P., Thomas-Hall S., Stephens E., Marx U., Mussgnug J., Posten C., Kruse O., Hankamer B. (2008) Second generation biofuels. High efficiency microalgae for biodiesel production. BioEnergy Res. 1, 20–43
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
