

Role for two conserved intermembrane space proteins, Ups1p and Ups2p, [corrected] in intra-mitochondrial phospholipid trafficking
- PMID: 22403410
- PMCID: PMC3346110
- DOI: 10.1074/jbc.M111.338665
Role for two conserved intermembrane space proteins, Ups1p and Ups2p, [corrected] in intra-mitochondrial phospholipid trafficking
Erratum in
- J Biol Chem. 2012 Aug 10;287(33):27450
Abstract
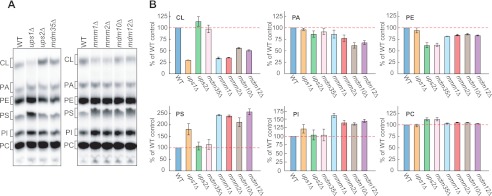
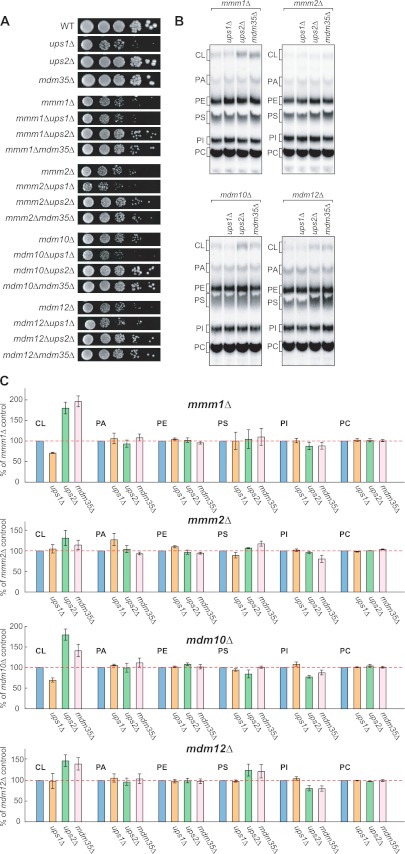
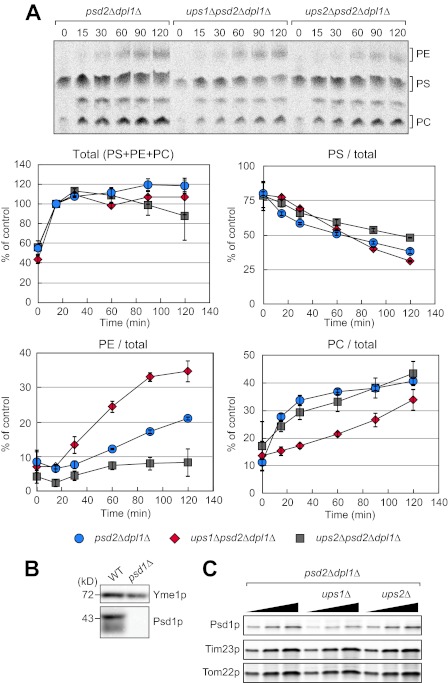
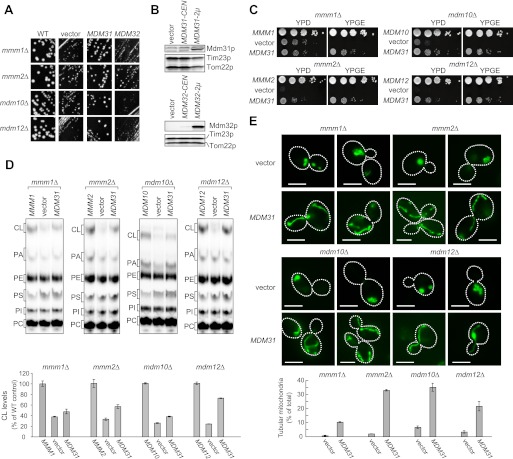
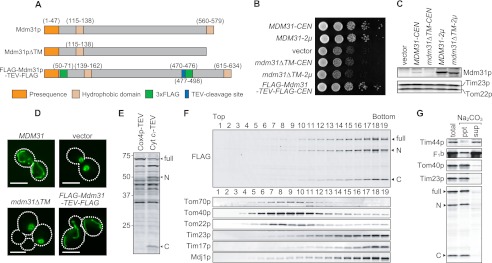
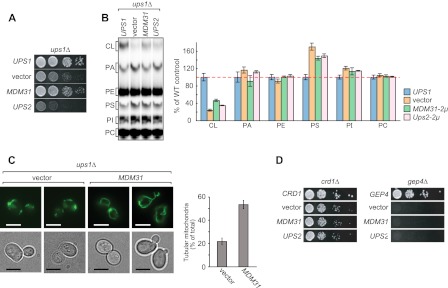
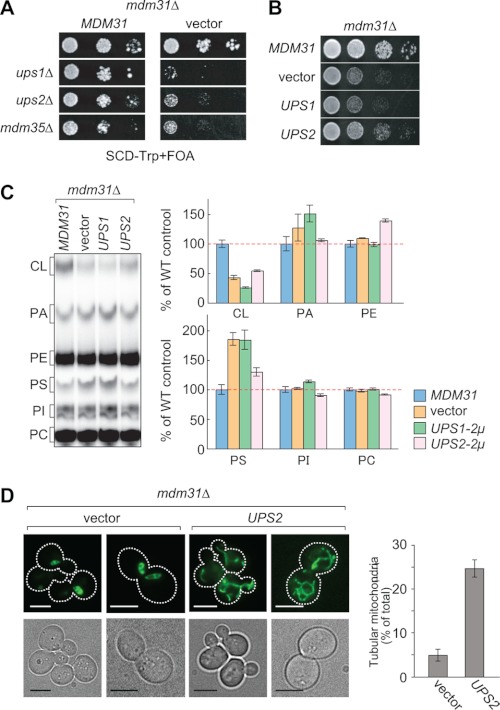
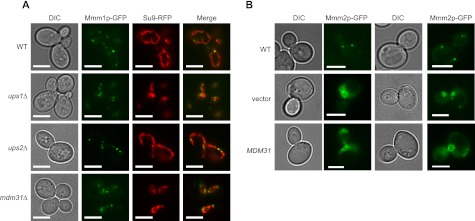
Mitochondrial membranes maintain a specific phospholipid composition. Most phospholipids are synthesized in the endoplasmic reticulum (ER) and transported to mitochondria, but cardiolipin and phosphatidylethanolamine are produced in mitochondria. In the yeast Saccharomyces cerevisiae, phospholipid exchange between the ER and mitochondria relies on the ER-mitochondria encounter structure (ERMES) complex, which physically connects the ER and mitochondrial outer membrane. However, the proteins and mechanisms involved in phospholipid transport within mitochondria remain elusive. Here, we investigated the role of the conserved intermembrane space proteins, Ups1p and Ups2p, and an inner membrane protein, Mdm31p, in phospholipid metabolism. Our data show that loss of the ERMES complex, Ups1p, and Mdm31p causes similar defects in mitochondrial phospholipid metabolism, mitochondrial morphology, and cell growth. Defects in cells lacking the ERMES complex or Ups1p are suppressed by Mdm31p overexpression as well as additional loss of Ups2p, which antagonizes Ups1p. Combined loss of the ERMES complex and Ups1p exacerbates phospholipid defects. Finally, pulse-chase experiments using [(14)C]serine revealed that Ups1p and Ups2p antagonistically regulate conversion of phosphatidylethanolamine to phosphatidylcholine. Our results suggest that Ups proteins and Mdm31p play important roles in phospholipid biosynthesis in mitochondria. Ups proteins may function in phospholipid trafficking between the outer and inner mitochondrial membranes.
Figures
References
-
- Jiang F., Ryan M. T., Schlame M., Zhao M., Gu Z., Klingenberg M., Pfanner N., Greenberg M. L. (2000) Absence of cardiolipin in the crd1 null mutant results in decreased mitochondrial membrane potential and reduced mitochondrial function. J. Biol. Chem. 275, 22387–22394 - PubMed
-
- van der Laan M., Meinecke M., Dudek J., Hutu D. P., Lind M., Perschil I., Guiard B., Wagner R., Pfanner N., Rehling P. (2007) Motor-free mitochondrial presequence translocase drives membrane integration of preproteins. Nat. Cell Biol. 9, 1152–1159 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
