

A Role for K2P Channels in the Operation of Somatosensory Nociceptors
- PMID: 22403526
- PMCID: PMC3293133
- DOI: 10.3389/fnmol.2012.00021
A Role for K2P Channels in the Operation of Somatosensory Nociceptors
Abstract
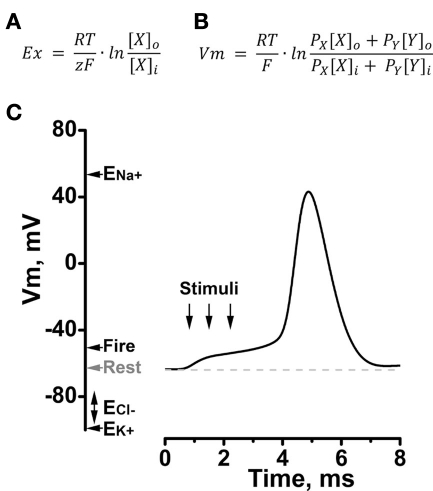
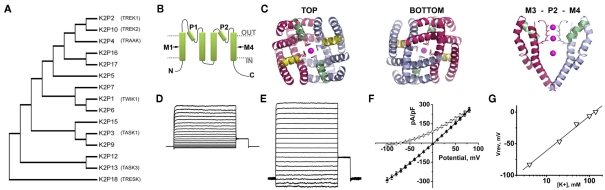
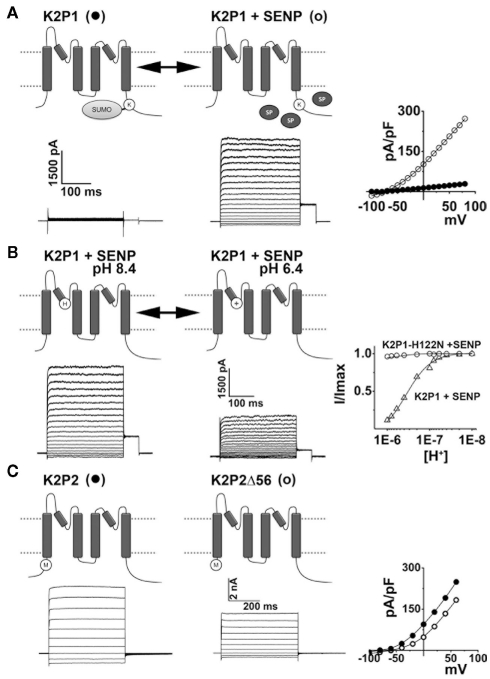
The ability to sense mechanical, thermal, and chemical stimuli is critical to normal physiology and the perception of pain. Contact with noxious stimuli triggers a complex series of events that initiate innate protective mechanisms designed to minimize or avoid injury. Extreme temperatures, mechanical stress, and chemical irritants are detected by specific ion channels and receptors clustered on the terminals of nociceptive sensory nerve fibers and transduced into electrical information. Propagation of these signals, from distant sites in the body to the spinal cord and the higher processing centers of the brain, is also orchestrated by distinct groups of ion channels. Since their identification in 1995, evidence has emerged to support roles for K2P channels at each step along this pathway, as receptors for physiological and noxious stimuli, and as determinants of nociceptor excitability and conductivity. In addition, the many subtypes of K2P channels expressed in somatosensory neurons are also implicated in mediating the effects of volatile, general anesthetics on the central and peripheral nervous systems. Here, I offer a critical review of the existing data supporting these attributes of K2P channel function and discuss how diverse regulatory mechanisms that control the activity of K2P channels act to govern the operation of nociceptors.
Keywords: K1P18; K2P1; K2P10; K2P2; K2P3; K2P4; K2P9; nociceptor.
Figures
References
-
- Alloui A., Zimmermann K., Mamet J., Duprat F., Noel J., Chemin J., Guy N., Blondeau N., Voilley N., Rubat-Coudert C., Borsotto M., Romey G., Heurteaux C., Reeh P., Eschalier A., Lazdunski M. (2006). TREK-1, a K+ channel involved in polymodal pain perception. EMBO J. 25, 2368–2376 10.1038/sj.emboj.7601116 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources