

Ontogeny of neural circuits underlying spatial memory in the rat
- PMID: 22403529
- PMCID: PMC3290765
- DOI: 10.3389/fncir.2012.00008
Ontogeny of neural circuits underlying spatial memory in the rat
Abstract
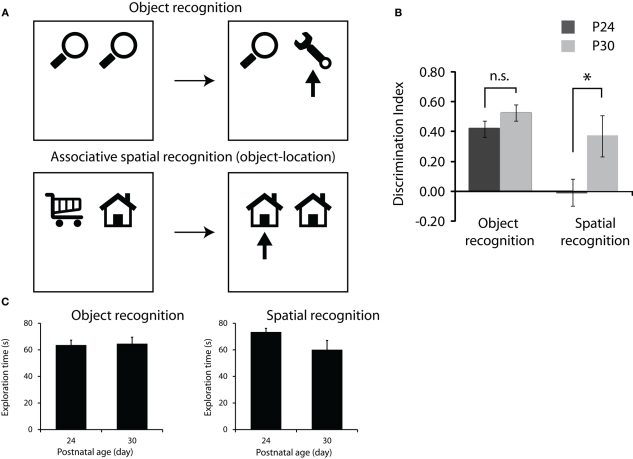
Spatial memory is a well-characterized psychological function in both humans and rodents. The combined computations of a network of systems including place cells in the hippocampus, grid cells in the medial entorhinal cortex and head direction cells found in numerous structures in the brain have been suggested to form the neural instantiation of the cognitive map as first described by Tolman in 1948. However, while our understanding of the neural mechanisms underlying spatial representations in adults is relatively sophisticated, we know substantially less about how this network develops in young animals. In this article we briefly review studies examining the developmental timescale that these systems follow. Electrophysiological recordings from very young rats show that directional information is at adult levels at the outset of navigational experience. The systems supporting allocentric memory, however, take longer to mature. This is consistent with behavioral studies of young rats which show that spatial memory based on head direction develops very early but that allocentric spatial memory takes longer to mature. We go on to report new data demonstrating that memory for associations between objects and their spatial locations is slower to develop than memory for objects alone. This is again consistent with previous reports suggesting that adult like spatial representations have a protracted development in rats and also suggests that the systems involved in processing non-spatial stimuli come online earlier.
Keywords: entorhinal cortex; grid cell; head direction cell; hippocampus; memory and learning; place cell; postnatal development; spatial representation.
Figures

References
LinkOut - more resources
Full Text Sources