

Bacterial mechanosensitive channels--MscS: evolution's solution to creating sensitivity in function
- PMID: 22404681
- PMCID: PMC3378650
- DOI: 10.1146/annurev-biophys-101211-113227
Bacterial mechanosensitive channels--MscS: evolution's solution to creating sensitivity in function
Abstract
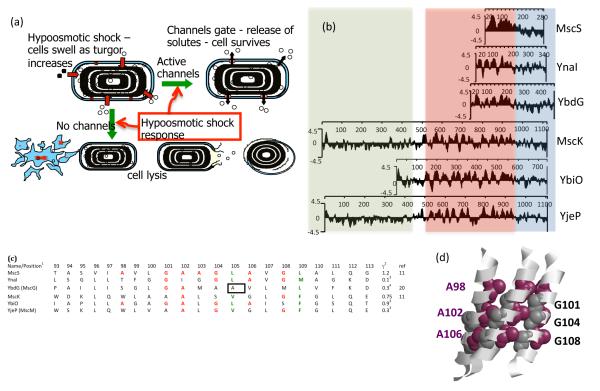
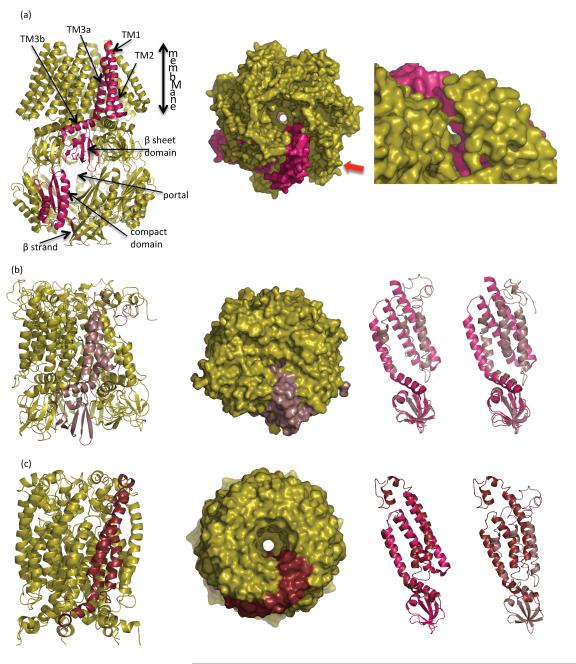
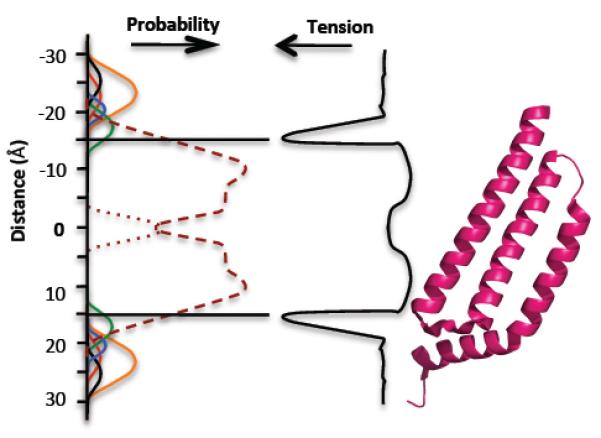
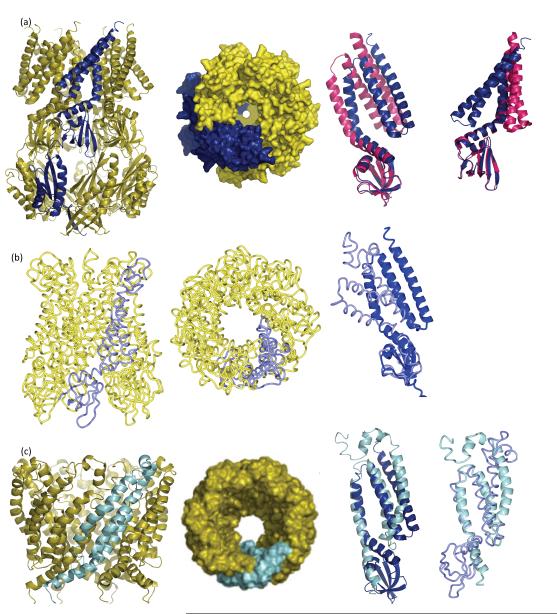
The discovery of mechanosensing channels has changed our understanding of bacterial physiology. The mechanosensitive channel of small conductance (MscS) is perhaps the most intensively studied of these channels. MscS has at least two states: closed, which does not allow solutes to exit the cytoplasm, and open, which allows rapid efflux of solvent and solutes. The ability to appropriately open or close the channel (gating) is critical to bacterial survival. We briefly review the science that led to the isolation and identification of MscS. We concentrate on the structure-function relationship of the channel, in particular the structural and biochemical approaches to understanding channel gating. We highlight the troubling discrepancies between the various models developed to understand MscS gating.
Figures
References
-
- Adamian L, Jackups R, Jr, Binkowski TA, Liang J. Higher-order interhelical spatial interactions in membrane proteins. J. Mol. Biol. 2003;327:251–72. - PubMed
-
- Adamian L, Liang J. Interhelical hydrogen bonds and spatial motifs in membrane proteins: polar clamps and serine zippers. Proteins. 2002;47:209–18. - PubMed
-
- Akitake B, Anishkin A, Liu N, Sukharev S. Straightening and sequential buckling of the pore-lining helices define the gating cycle of MscS. Nat. Struct. Mol. Biol. 2007;14:1141–49. - PubMed
-
- Reports extended dynamics analysis of MscS in both open and closed forms and focuses on the kinking and unkinking within TM3b.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
