

Single-molecule views of protein movement on single-stranded DNA
- PMID: 22404684
- PMCID: PMC3719979
- DOI: 10.1146/annurev-biophys-042910-155351
Single-molecule views of protein movement on single-stranded DNA
Abstract
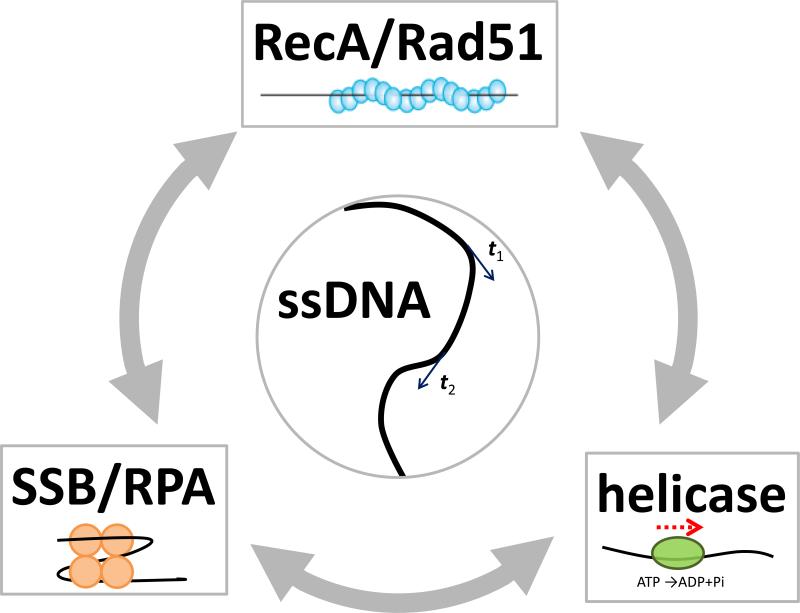
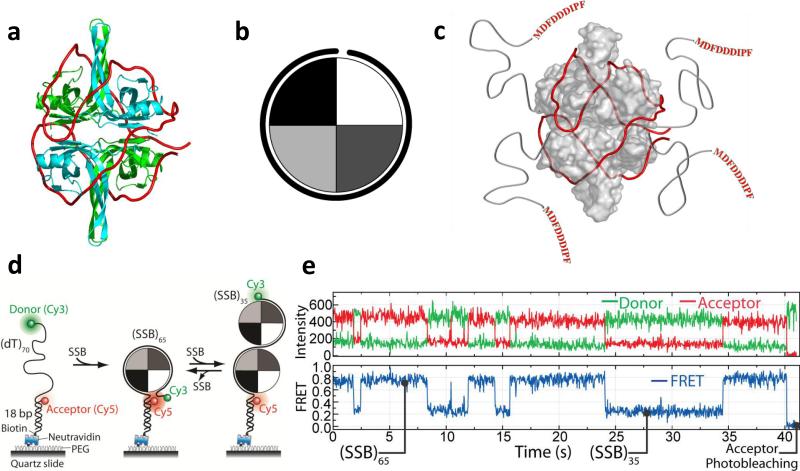
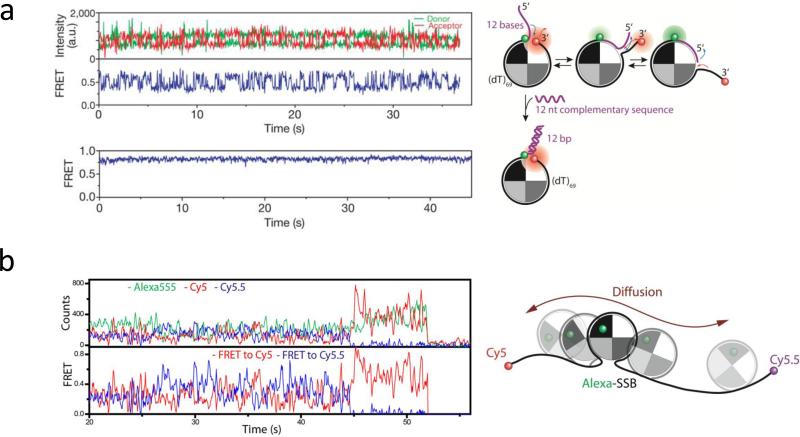
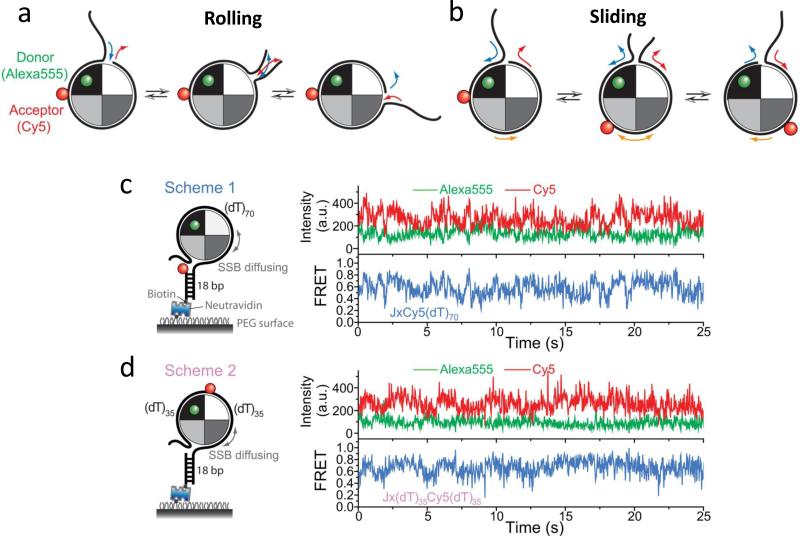
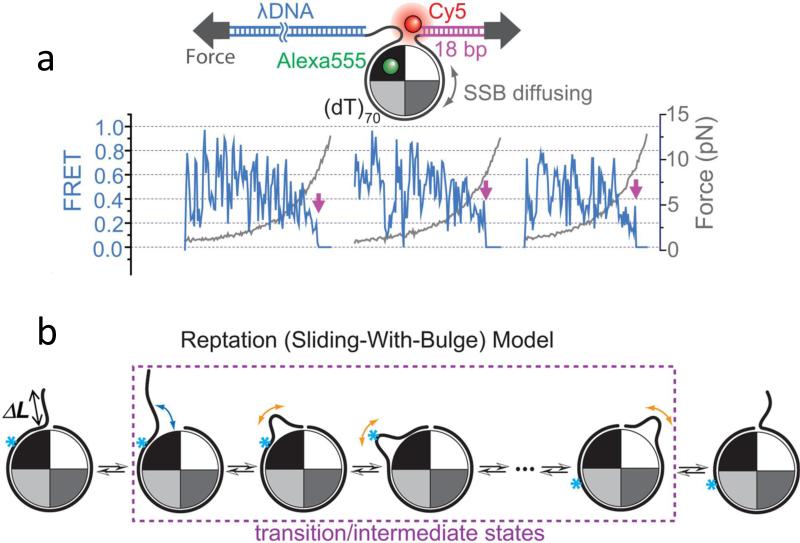
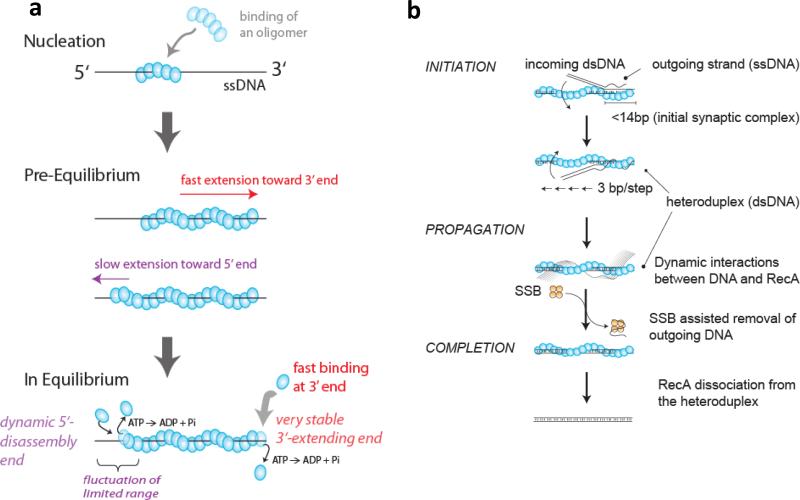
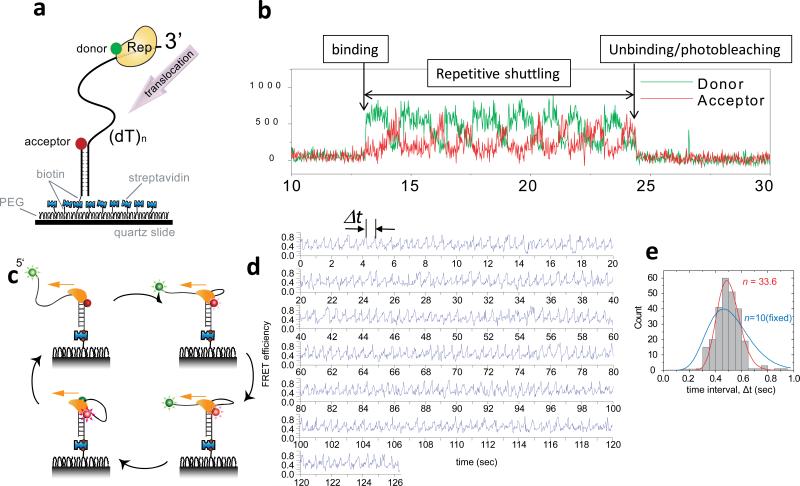
The advent of new technologies allowing the study of single biological molecules continues to have a major impact on studies of interacting systems as well as enzyme reactions. These approaches (fluorescence, optical, and magnetic tweezers), in combination with ensemble methods, have been particularly useful for mechanistic studies of protein-nucleic acid interactions and enzymes that function on nucleic acids. We review progress in the use of single-molecule methods to observe and perturb the activities of proteins and enzymes that function on flexible single-stranded DNA. These include single-stranded DNA binding proteins, recombinases (RecA/Rad51), and helicases/translocases that operate as motor proteins and play central roles in genome maintenance. We emphasize methods that have been used to detect and study the movement of these proteins (both ATP-dependent directional and random movement) along the single-stranded DNA and the mechanistic and functional information that can result from detailed analysis of such movement.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
