

Fumarate is cardioprotective via activation of the Nrf2 antioxidant pathway
- PMID: 22405071
- PMCID: PMC3314920
- DOI: 10.1016/j.cmet.2012.01.017
Fumarate is cardioprotective via activation of the Nrf2 antioxidant pathway
Abstract
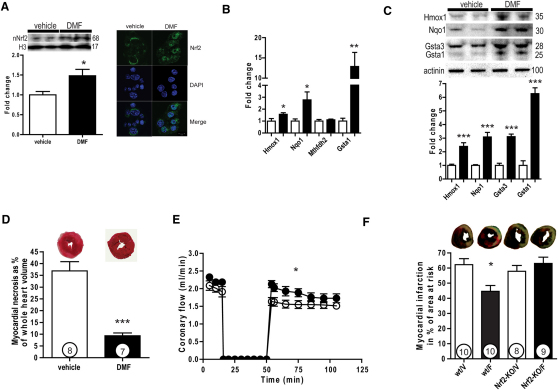
The citric acid cycle (CAC) metabolite fumarate has been proposed to be cardioprotective; however, its mechanisms of action remain to be determined. To augment cardiac fumarate levels and to assess fumarate's cardioprotective properties, we generated fumarate hydratase (Fh1) cardiac knockout (KO) mice. These fumarate-replete hearts were robustly protected from ischemia-reperfusion injury (I/R). To compensate for the loss of Fh1 activity, KO hearts maintain ATP levels in part by channeling amino acids into the CAC. In addition, by stabilizing the transcriptional regulator Nrf2, Fh1 KO hearts upregulate protective antioxidant response element genes. Supporting the importance of the latter mechanism, clinically relevant doses of dimethylfumarate upregulated Nrf2 and its target genes, hence protecting control hearts, but failed to similarly protect Nrf2-KO hearts in an in vivo model of myocardial infarction. We propose that clinically established fumarate derivatives activate the Nrf2 pathway and are readily testable cytoprotective agents.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Adam J., Hatipoglu E., O'Flaherty L., Ternette N., Sahgal N., Lockstone H., Baban D., Nye E., Stamp G.W., Wolhuter K. Renal cyst formation in Fh1-deficient mice is independent of the Hif/Phd pathway: roles for fumarate in KEAP1 succination and Nrf2 signaling. Cancer Cell. 2011;20:524–537. - PMC - PubMed
-
- Adams C.M. Role of the transcription factor ATF4 in the anabolic actions of insulin and the anti-anabolic actions of glucocorticoids. J. Biol. Chem. 2007;282:16744–16753. - PubMed
-
- Afonyushkin T., Oskolkova O.V., Philippova M., Resink T.J., Erne P., Binder B.R., Bochkov V.N. Oxidized phospholipids regulate expression of ATF4 and VEGF in endothelial cells via NRF2-dependent mechanism: novel point of convergence between electrophilic and unfolded protein stress pathways. Arterioscler. Thromb. Vasc. Biol. 2010;30:1007–1013. - PubMed
-
- Appleton S.D., Chretien M.L., McLaughlin B.E., Vreman H.J., Stevenson D.K., Brien J.F., Nakatsu K., Maurice D.H., Marks G.S. Selective inhibition of heme oxygenase, without inhibition of nitric oxide synthase or soluble guanylyl cyclase, by metalloporphyrins at low concentrations. Drug Metab. Dispos. 1999;27:1214–1219. - PubMed
-
- Ashrafian H., O'Flaherty L., Adam J., Steeples V., Chung Y.L., East P., Vanharanta S., Lehtonen H., Nye E., Hatipoglu E. Expression profiling in progressive stages of fumarate-hydratase deficiency: the contribution of metabolic changes to tumorigenesis. Cancer Res. 2010;70:9153–9165. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
