

Checkpoint-independent stabilization of kinetochore-microtubule attachments by Mad2 in human cells
- PMID: 22405866
- PMCID: PMC3326208
- DOI: 10.1016/j.cub.2012.02.030
Checkpoint-independent stabilization of kinetochore-microtubule attachments by Mad2 in human cells
Abstract
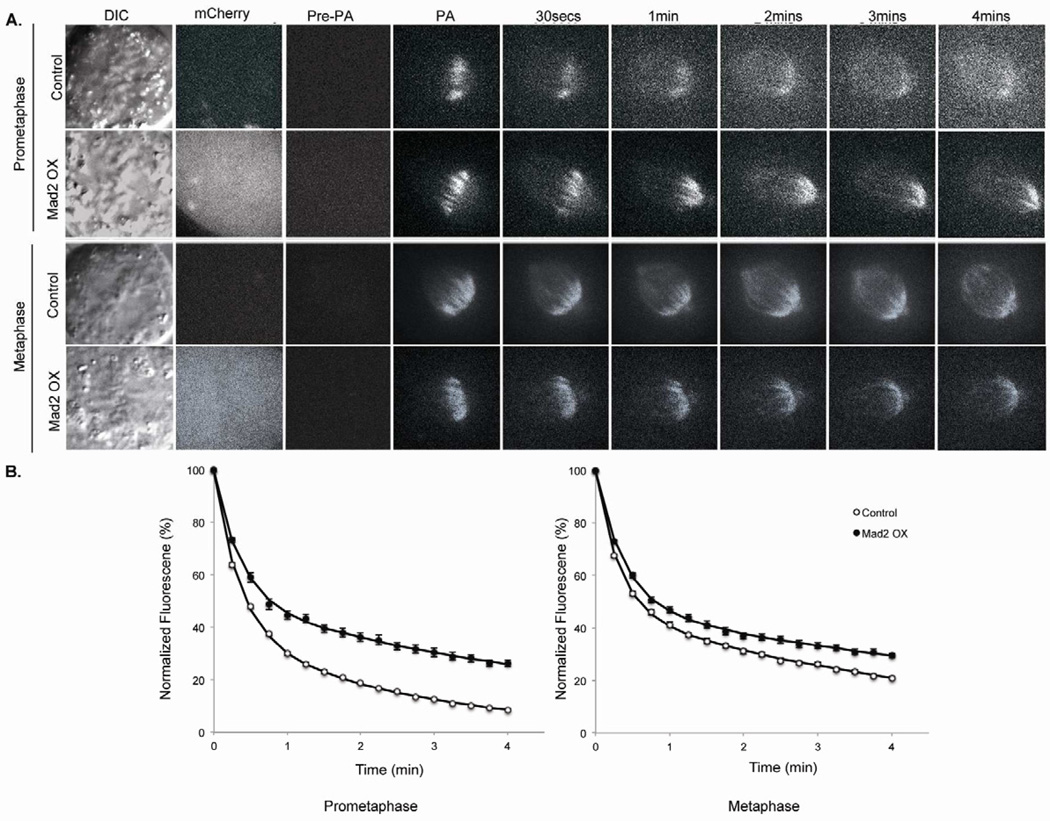
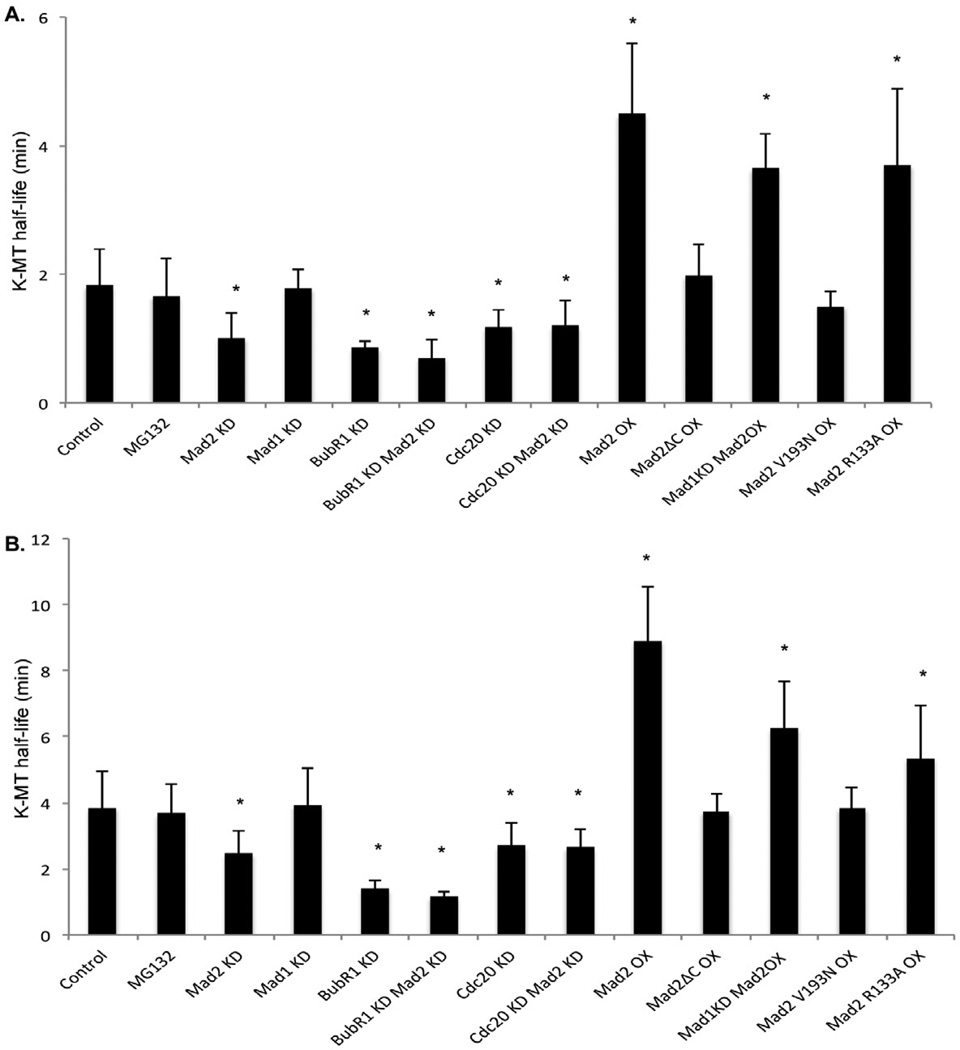
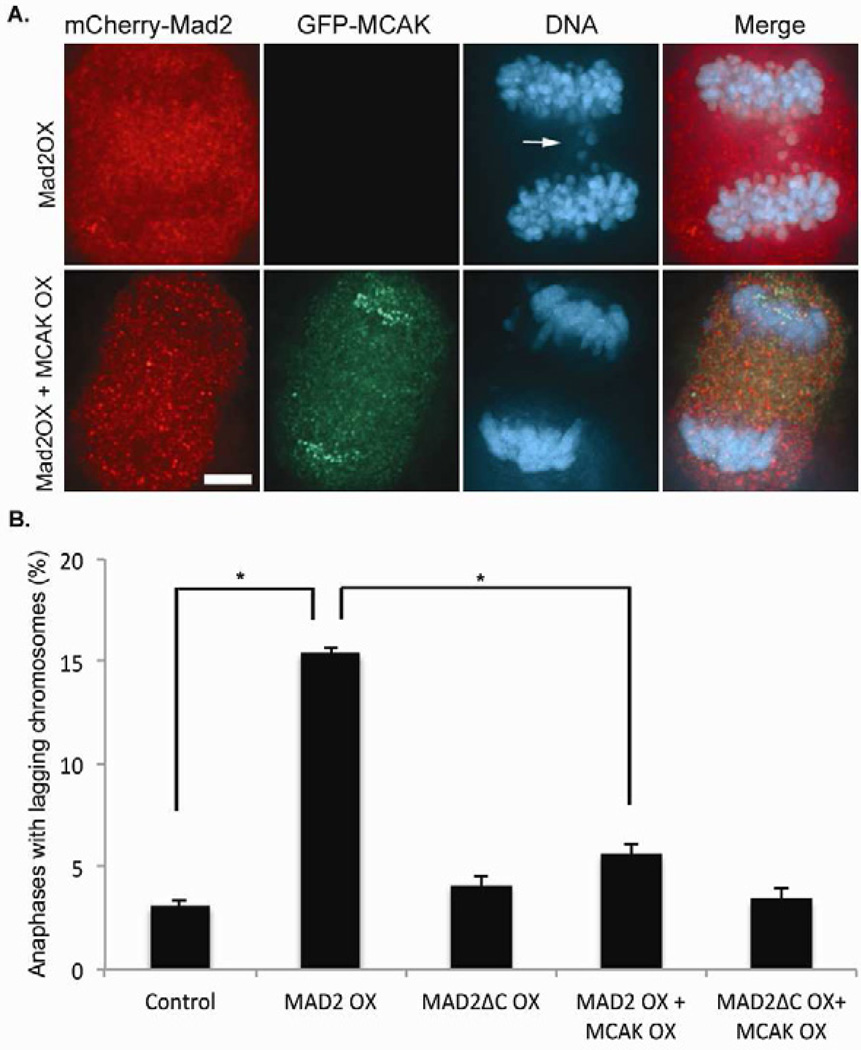
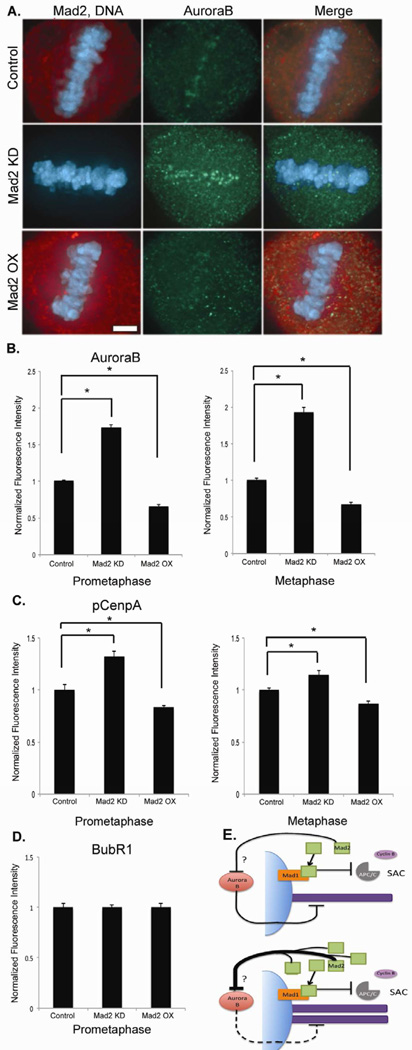
Faithful chromosome segregation is required for cell and organism viability and relies on both the mitotic checkpoint and the machinery that corrects kinetochore-microtubule (k-MT) attachment errors. Most solid tumors have aneuploid karyotypes and many missegregate chromosomes at high rates in a phenomenon called chromosomal instability (CIN). Mad2 is essential for mitotic checkpoint function and is frequently overexpressed in human tumors that are CIN. For unknown reasons, cells overexpressing Mad2 display high rates of lagging chromosomes. Here, we explore this phenomenon and show that k-MT attachments are hyperstabilized by Mad2 overexpression and that this undermines the efficiency of correction of k-MT attachment errors. Mad2 affects k-MT attachment stability independently of the mitotic checkpoint because k-MT attachments are unaltered upon Mad1 depletion and Mad2 overexpression hyperstabilizes k-MT attachments in Mad1-deficient cells. Mad2 mediates these effects with Cdc20 by altering the centromeric localization and activity of Aurora B kinase, a known regulator of k-MT attachment stability. These data reveal a new function for Mad2 to stabilize k-MT attachments independent of the checkpoint and explain why Mad2 overexpression increases chromosome missegregation to cause chromosomal instability in human tumors.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Chromosomal Instability: Mad2 beyond the spindle checkpoint.Curr Biol. 2012 Apr 10;22(7):R233-5. doi: 10.1016/j.cub.2012.02.059. Curr Biol. 2012. PMID: 22497940
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
