

Structural, functional and evolutionary relationships between homing endonucleases and proteins from their host organisms
- PMID: 22406833
- PMCID: PMC3384342
- DOI: 10.1093/nar/gks226
Structural, functional and evolutionary relationships between homing endonucleases and proteins from their host organisms
Abstract
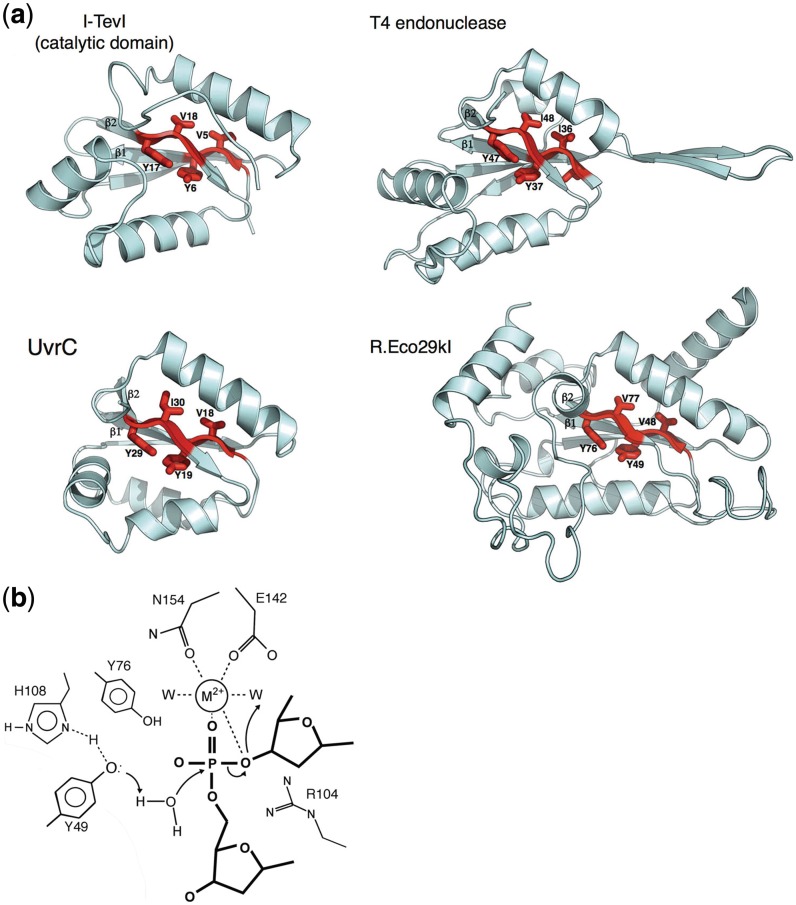
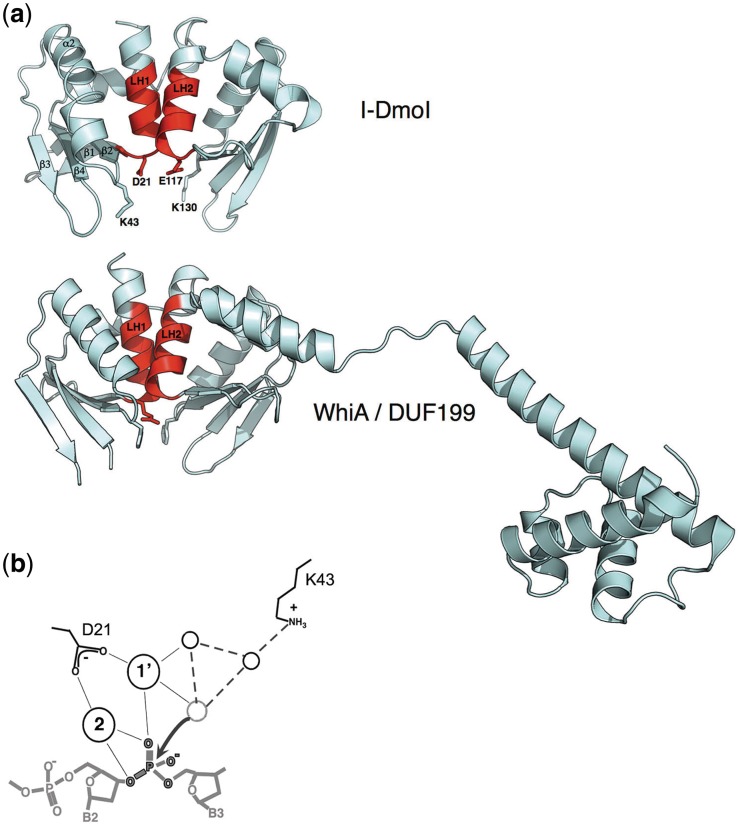
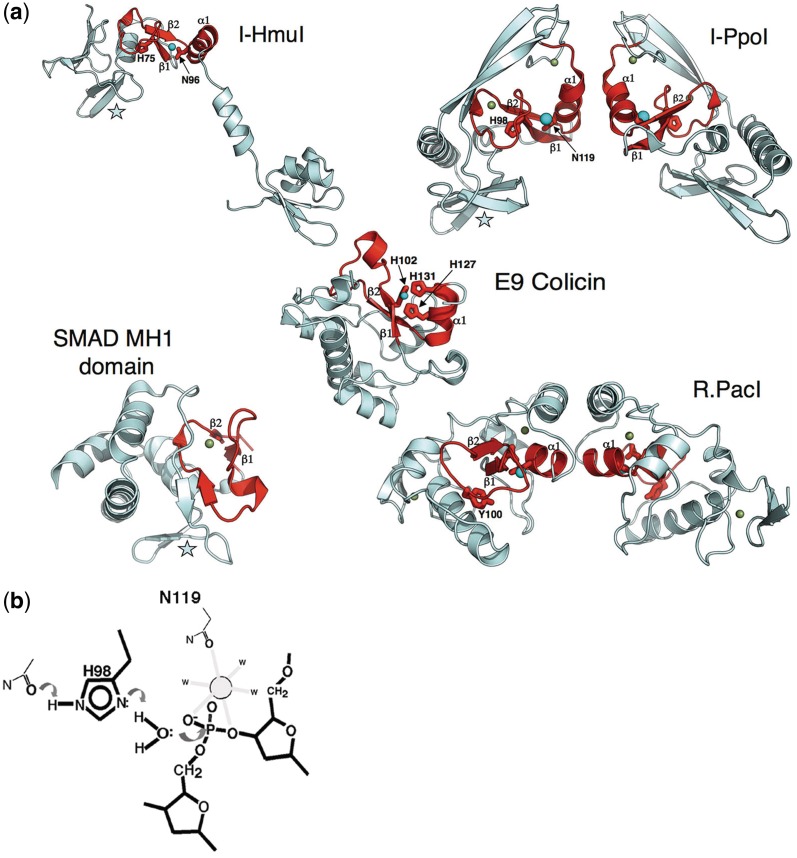
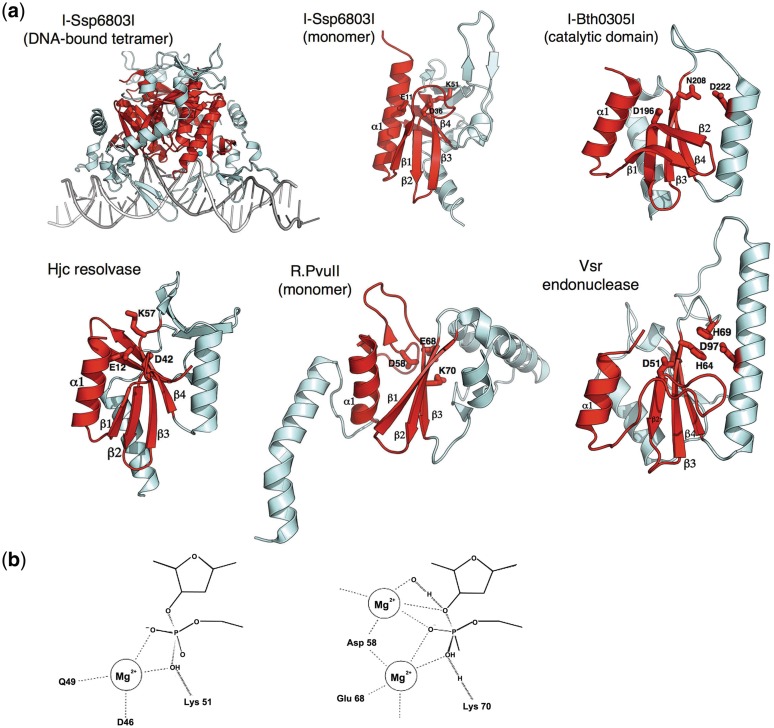
Homing endonucleases (HEs) are highly specific DNA-cleaving enzymes that are encoded by invasive DNA elements (usually mobile introns or inteins) within the genomes of phage, bacteria, archea, protista and eukaryotic organelles. Six unique structural HE families, that collectively span four distinct nuclease catalytic motifs, have been characterized to date. Members of each family display structural homology and functional relationships to a wide variety of proteins from various organisms. The biological functions of those proteins are highly disparate and include non-specific DNA-degradation enzymes, restriction endonucleases, DNA-repair enzymes, resolvases, intron splicing factors and transcription factors. These relationships suggest that modern day HEs share common ancestors with proteins involved in genome fidelity, maintenance and gene expression. This review summarizes the results of structural studies of HEs and corresponding proteins from host organisms that have illustrated the manner in which these factors are related.
Figures
References
-
- Dujon B. Group I introns as mobile genetic elements: facts and mechanistic speculations – a review. Gene. 1989;82:91–114. - PubMed
-
- Dalgaard JZ, Klar AJ, Moser MJ, Holley WR, Chatterjee A, Mian IS. Statistical modeling and analysis of the LAGLIDADG family of site-specific endonucleases and identification of an intein that encodes a site-specific endonuclease of the HNH family. Nucleic Acids Res. 1997;25:4626–4638. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
