

Calpain-mediated vimentin cleavage occurs upstream of MT1-MMP membrane translocation to facilitate endothelial sprout initiation
- PMID: 22407449
- PMCID: PMC3338915
- DOI: 10.1007/s10456-012-9262-4
Calpain-mediated vimentin cleavage occurs upstream of MT1-MMP membrane translocation to facilitate endothelial sprout initiation
Abstract
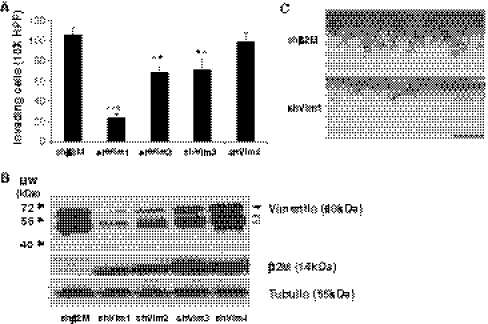
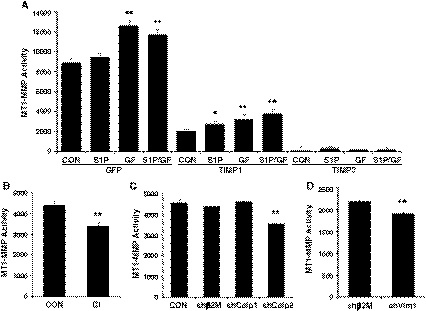
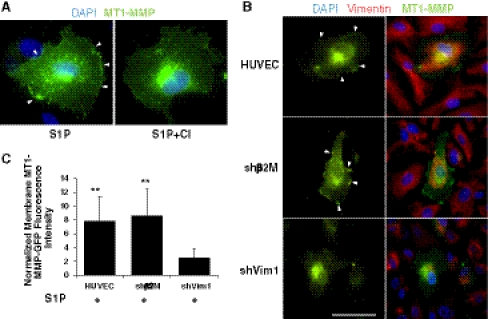
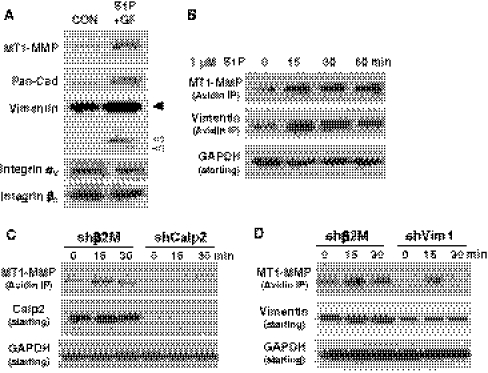
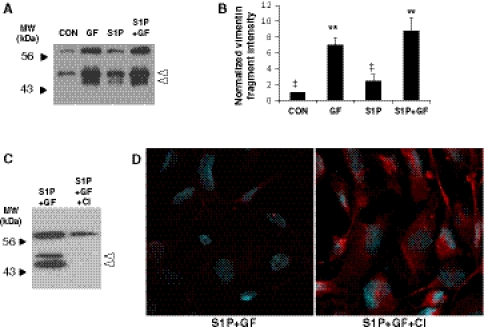
Endothelial cells normally line the vasculature and remain quiescent. However, these cells can be rapidly stimulated to undergo morphogenesis and initiate new blood vessel formation given the proper cues. This study reports a new mechanism for initiating angiogenic sprout formation that involves vimentin, the major intermediate filament protein in endothelial cells. Initial studies confirmed vimentin was required for sphingosine 1-phosphate (S1P)- and growth factor (GF)-induced endothelial cell invasion, and vimentin was cleaved by calpains during invasion. Calpains were predominantly activated by GF and were required for sprout initiation. Because others have reported membrane type 1-matrix metalloproteinase (MT1-MMP) is required for endothelial sprouting responses, we tested whether vimentin and calpain acted upstream of MT1-MMP. Both calpain and vimentin were required for successful MT1-MMP membrane translocation, which was stimulated by S1P. In addition, vimentin complexed with MT1-MMP in a manner that required both the cytoplasmic domain of MT1-MMP and calpain activation, which increased the soluble pool of vimentin in endothelial cells. Altogether, these data indicate that pro-angiogenic signals converge to activate calpain-dependent vimentin cleavage and increase vimentin solubility, which act upstream to facilitate MT1-MMP membrane translocation, resulting in successful endothelial sprout formation in three-dimensional collagen matrices. These findings help explain why S1P and GF synergize to stimulate robust sprouting in 3D collagen matrices.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
