

CEF1/CDC5 alleles modulate transitions between catalytic conformations of the spliceosome
- PMID: 22408182
- PMCID: PMC3334688
- DOI: 10.1261/rna.029421.111
CEF1/CDC5 alleles modulate transitions between catalytic conformations of the spliceosome
Abstract
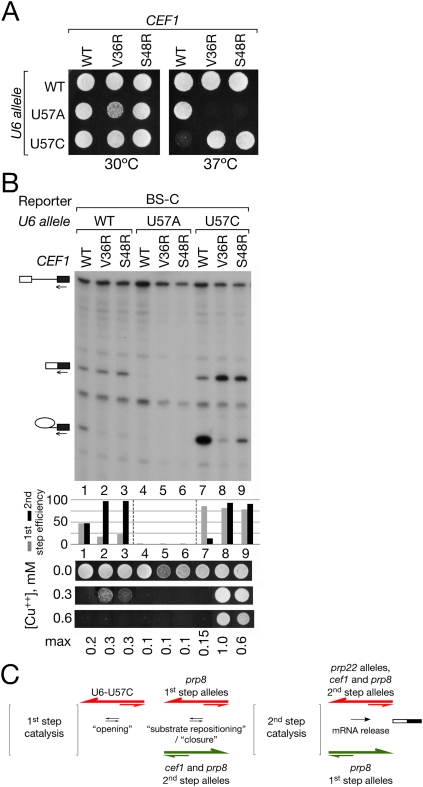
Conformational change within the spliceosome is required between the first and second catalytic steps of pre-mRNA splicing. A prior genetic screen for suppressors of an intron mutant that stalls between the two steps yielded both prp8 and non-prp8 alleles that suppressed second-step splicing defects. We have now identified the strongest non-prp8 suppressors as alleles of the NTC (Prp19 complex) component, CEF1. These cef1 alleles generally suppress second-step defects caused by a variety of intron mutations, mutations in U6 snRNA, or deletion of the second-step protein factor Prp17, and they can activate alternative 3' splice sites. Genetic and functional interactions between cef1 and prp8 alleles suggest that they modulate the same event(s) in the first-to-second-step transition, most likely by stabilization of the second-step spliceosome; in contrast, alleles of U6 snRNA that also alter this transition modulate a distinct event, most likely by stabilization of the first-step spliceosome. These results implicate a myb-like domain of Cef1/CDC5 in interactions that modulate conformational states of the spliceosome and suggest that alteration of these events affects splice site use, resulting in alternative splicing-like patterns in yeast.
Figures






References
-
- Bessonov S, Anokhina M, Will CL, Urlaub H, Lührmann R 2008. Isolation of an active step I spliceosome and composition of its RNP core. Nature 452: 846–850 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous