

Betalain production is possible in anthocyanin-producing plant species given the presence of DOPA-dioxygenase and L-DOPA
- PMID: 22409631
- PMCID: PMC3317834
- DOI: 10.1186/1471-2229-12-34
Betalain production is possible in anthocyanin-producing plant species given the presence of DOPA-dioxygenase and L-DOPA
Abstract
Background: Carotenoids and anthocyanins are the predominant non-chlorophyll pigments in plants. However, certain families within the order Caryophyllales produce another class of pigments, the betalains, instead of anthocyanins. The occurrence of betalains and anthocyanins is mutually exclusive. Betalains are divided into two classes, the betaxanthins and betacyanins, which produce yellow to orange or violet colours, respectively. In this article we show betalain production in species that normally produce anthocyanins, through a combination of genetic modification and substrate feeding.
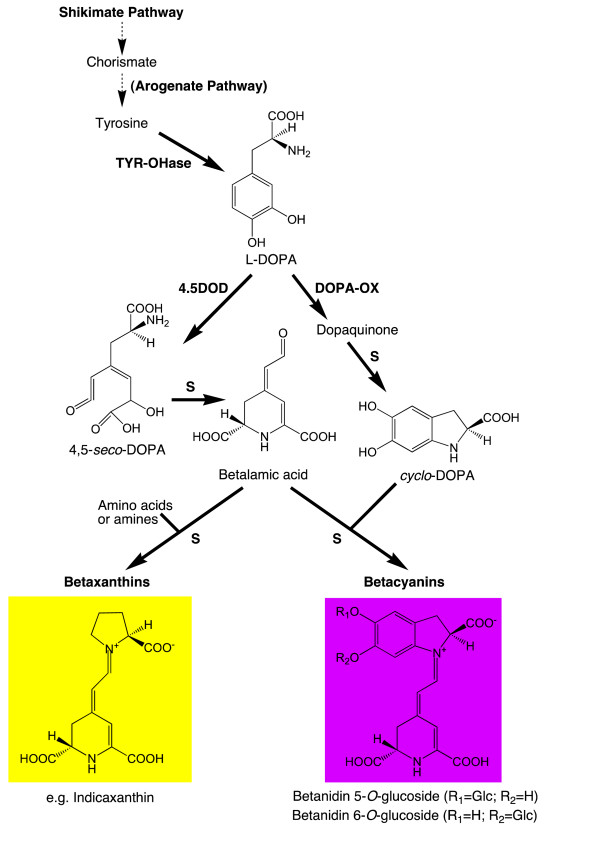
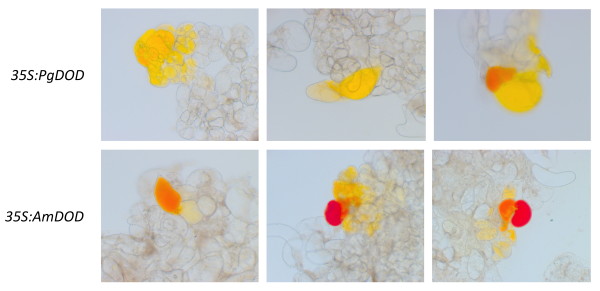
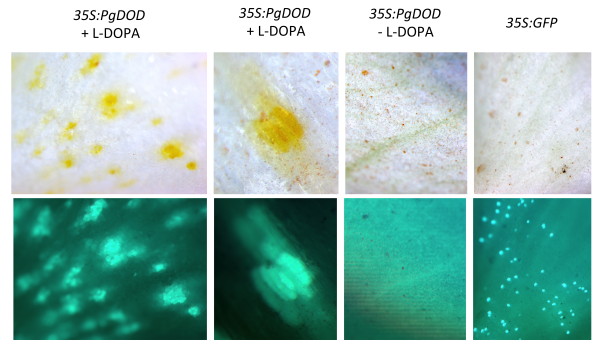
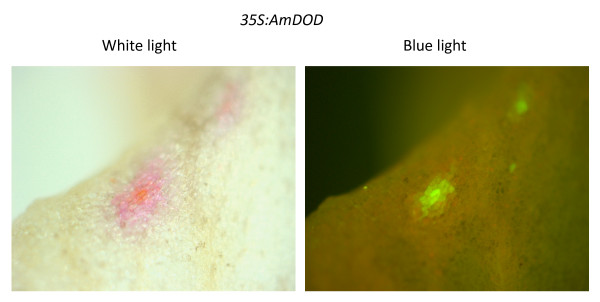
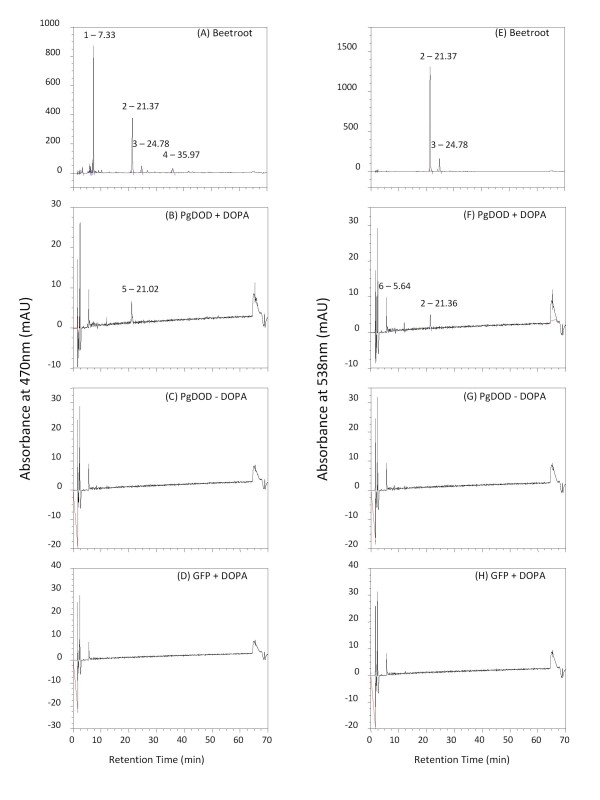
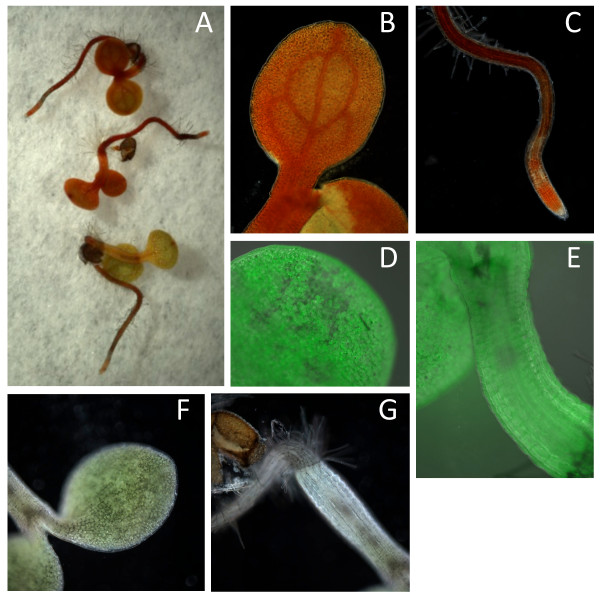
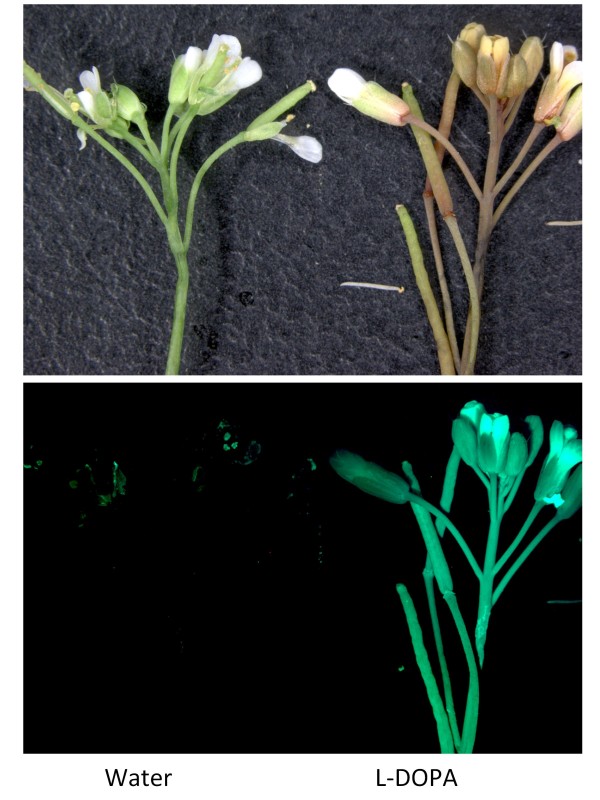
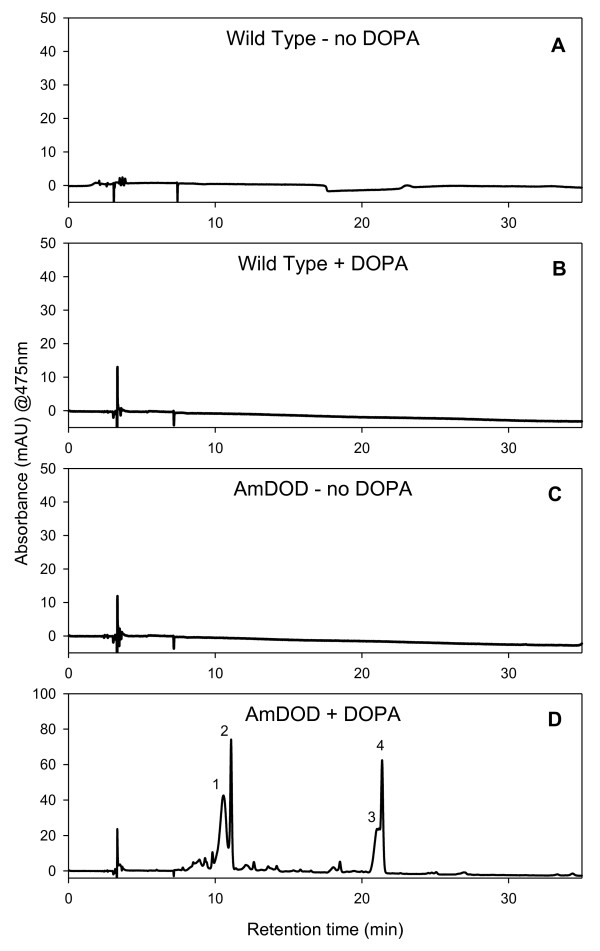
Results: The biolistic introduction of DNA constructs for transient overexpression of two different dihydroxyphenylalanine (DOPA) dioxygenases (DODs), and feeding of DOD substrate (L-DOPA), was sufficient to induce betalain production in cell cultures of Solanum tuberosum (potato) and petals of Antirrhinum majus. HPLC analysis showed both betaxanthins and betacyanins were produced. Multi-cell foci with yellow, orange and/or red colours occurred, with either a fungal DOD (from Amanita muscaria) or a plant DOD (from Portulaca grandiflora), and the yellow/orange foci showed green autofluorescence characteristic of betaxanthins. Stably transformed Arabidopsis thaliana (arabidopsis) lines containing 35S: AmDOD produced yellow colouration in flowers and orange-red colouration in seedlings when fed L-DOPA. These tissues also showed green autofluorescence. HPLC analysis of the transgenic seedlings fed L-DOPA confirmed betaxanthin production.
Conclusions: The fact that the introduction of DOD along with a supply of its substrate (L-DOPA) was sufficient to induce betacyanin production reveals the presence of a background enzyme, possibly a tyrosinase, that can convert L-DOPA to cyclo-DOPA (or dopaxanthin to betacyanin) in at least some anthocyanin-producing plants. The plants also demonstrate that betalains can accumulate in anthocyanin-producing species. Thus, introduction of a DOD and an enzyme capable of converting tyrosine to L-DOPA should be sufficient to confer both betaxanthin and betacyanin production to anthocyanin-producing species. The requirement for few novel biosynthetic steps may have assisted in the evolution of the betalain biosynthetic pathway in the Caryophyllales, and facilitated multiple origins of the pathway in this order and in fungi. The stably transformed 35S: AmDOD arabidopsis plants provide material to study, for the first time, the physiological effects of having both betalains and anthocyanins in the same plant tissues.
Figures
References
-
- Gould KS, Lister C. In: Flavonoids: Chemistry, Biochemistry and Applications. Andersen ØM, Markham KR, editor. CRC Press, Boca Raton; 2006. Flavonoid functions in plants; pp. 397–441.
-
- Davies KM. In: Plant Pigments and their Manipulation. Davies KM, editor. Blackwell Publishing, Oxford; 2004. An introduction to plant pigments in biology and commerce; pp. 1–22.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
