

Membrane vesicle release in bacteria, eukaryotes, and archaea: a conserved yet underappreciated aspect of microbial life
- PMID: 22409932
- PMCID: PMC3370574
- DOI: 10.1128/IAI.06014-11
Membrane vesicle release in bacteria, eukaryotes, and archaea: a conserved yet underappreciated aspect of microbial life
Abstract
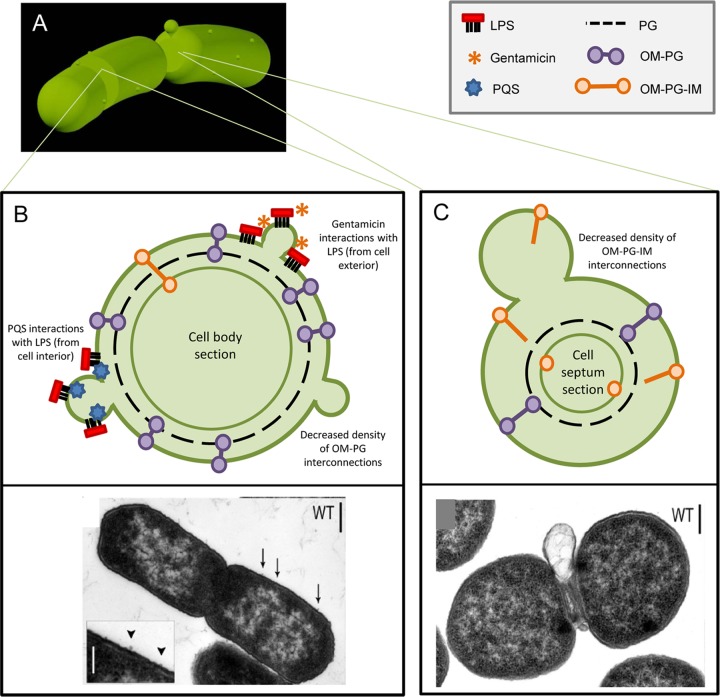
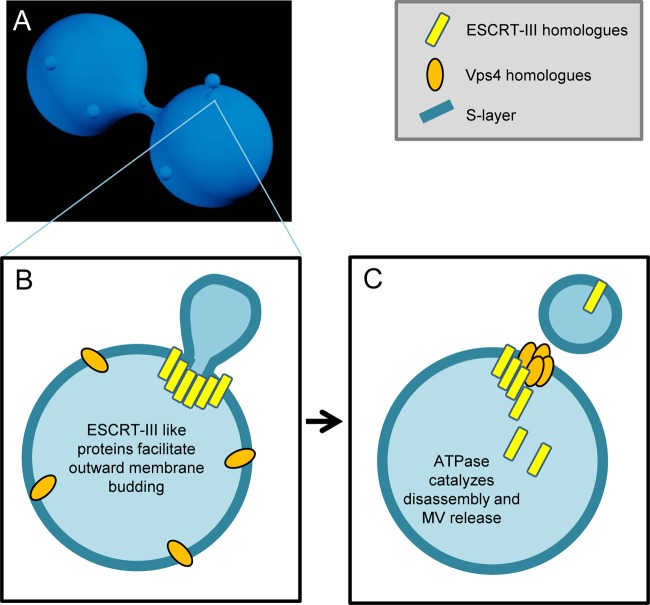
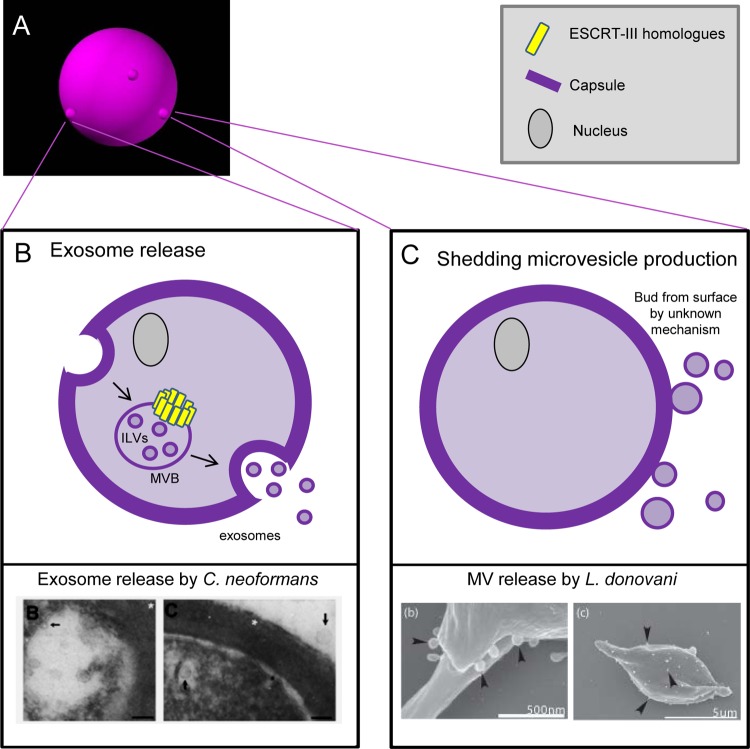
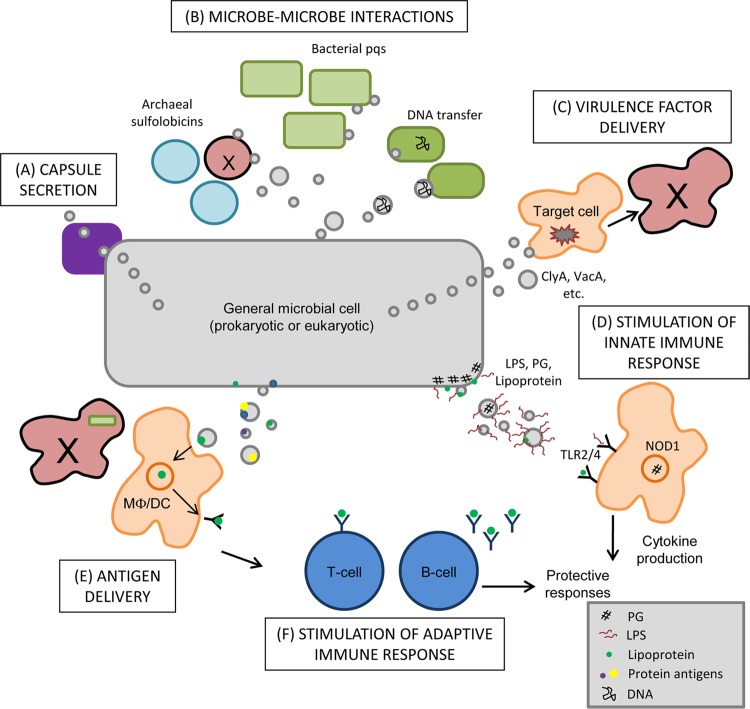
Interaction of microbes with their environment depends on features of the dynamic microbial surface throughout cell growth and division. Surface modifications, whether used to acquire nutrients, defend against other microbes, or resist the pressures of a host immune system, facilitate adaptation to unique surroundings. The release of bioactive membrane vesicles (MVs) from the cell surface is conserved across microbial life, in bacteria, archaea, fungi, and parasites. MV production occurs not only in vitro but also in vivo during infection, underscoring the influence of these surface organelles in microbial physiology and pathogenesis through delivery of enzymes, toxins, communication signals, and antigens recognized by the innate and adaptive immune systems. Derived from a variety of organisms that span kingdoms of life and called by several names (membrane vesicles, outer membrane vesicles [OMVs], exosomes, shedding microvesicles, etc.), the conserved functions and mechanistic strategies of MV release are similar, including the use of ESCRT proteins and ESCRT protein homologues to facilitate these processes in archaea and eukaryotic microbes. Although forms of MV release by different organisms share similar visual, mechanistic, and functional features, there has been little comparison across microbial life. This underappreciated conservation of vesicle release, and the resulting functional impact throughout the tree of life, explored in this review, stresses the importance of vesicle-mediated processes throughout biology.
Figures
References
-
- Abadi J, Pirofski L. 1999. Antibodies reactive with the cryptococcal capsular polysaccharide glucuronoxylomannan are present in sera from children with and without human immunodeficiency virus infection. J. Infect. Dis. 180:915–919 - PubMed
-
- Alaniz RC, Deatherage BL, Lara JC, Cookson BT. 2007. Membrane vesicles are immunogenic facsimiles of Salmonella typhimurium that potently activate dendritic cells, prime B and T cell responses, and stimulate protective immunity in vivo. J. Immunol. 179:7692–7701 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
