

Molecular underpinnings of centromere identity and maintenance
- PMID: 22410197
- PMCID: PMC3614489
- DOI: 10.1016/j.tibs.2012.01.003
Molecular underpinnings of centromere identity and maintenance
Abstract
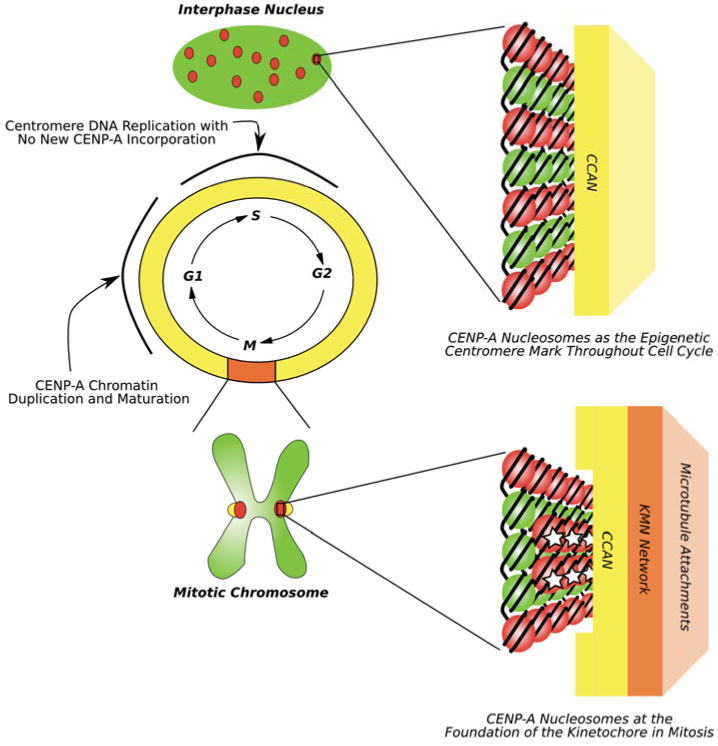
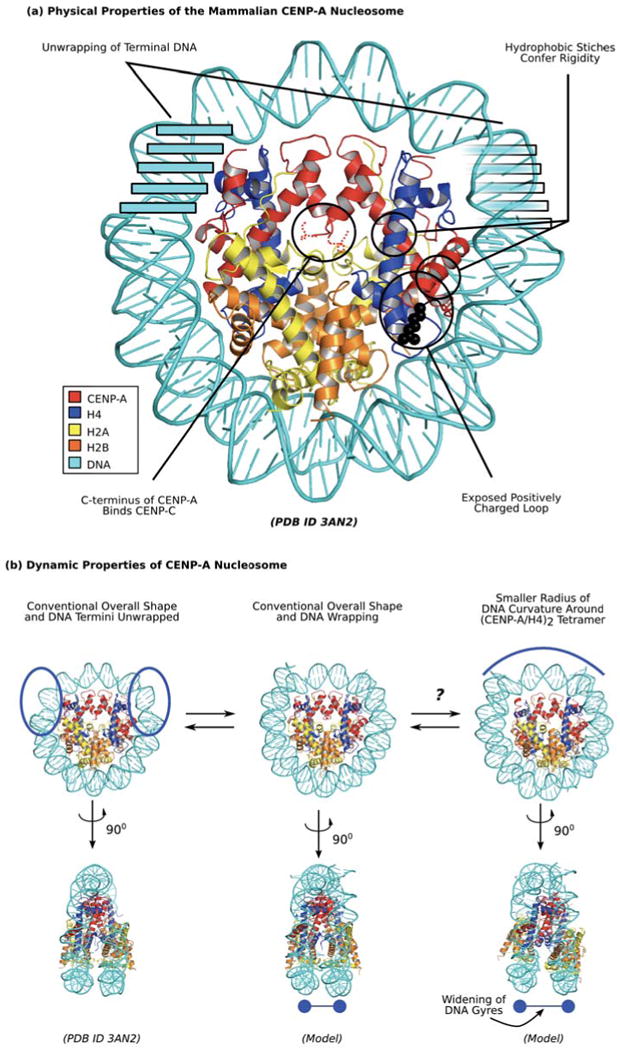
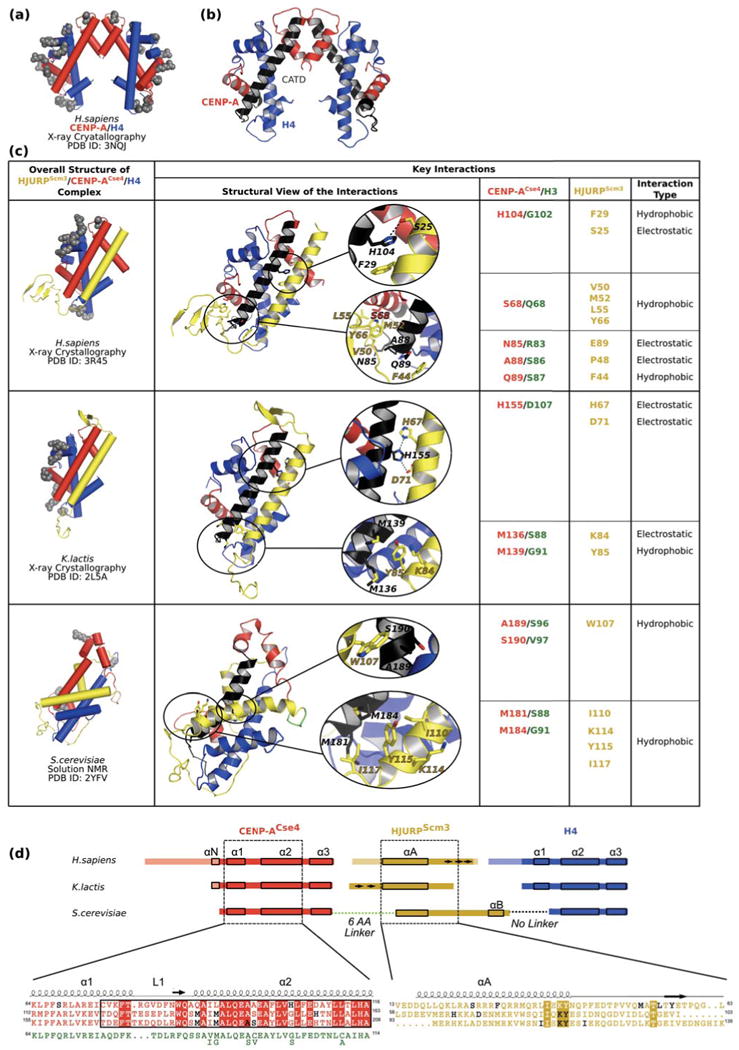
Centromeres direct faithful chromosome inheritance at cell division but are not defined by a conserved DNA sequence. Instead, a specialized form of chromatin containing the histone H3 variant, CENP-A, epigenetically specifies centromere location. We discuss current models where CENP-A serves as the marker for the centromere during the entire cell cycle in addition to generating the foundational chromatin for the kinetochore in mitosis. Recent elegant experiments have indicated that engineered arrays of CENP-A-containing nucleosomes are sufficient to serve as the site of kinetochore formation and for seeding centromeric chromatin that self-propagates through cell generations. Finally, recent structural and dynamic studies of CENP-A-containing histone complexes - before and after assembly into nucleosomes - provide models to explain underlying molecular mechanisms at the centromere.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures

References
-
- Cleveland DW, et al. Centromeres and kinetochores: from epigenetics to mitotic checkpoint signaling. Cell. 2003;112:407–421. - PubMed
-
- Eichler EE. Repetitive conundrums of centromere structure and function. Hum Mol Genet. 1999;8:151–155. - PubMed
-
- Henikoff S, et al. The centromere paradox: stable inheritance with rapidly evolving DNA. Science. 2001;293:1098–1102. - PubMed
-
- Murphy WJ, et al. Dynamics of mammalian chromosome evolution inferred from multispecies comparative maps. Science. 2005;309:613–617. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
