

Commensal bacteria modulate innate immune responses of vaginal epithelial cell multilayer cultures
- PMID: 22412914
- PMCID: PMC3296736
- DOI: 10.1371/journal.pone.0032728
Commensal bacteria modulate innate immune responses of vaginal epithelial cell multilayer cultures
Abstract
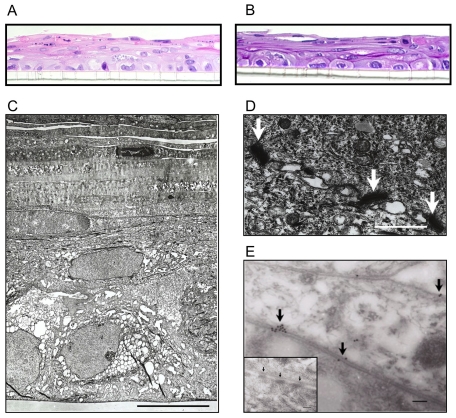
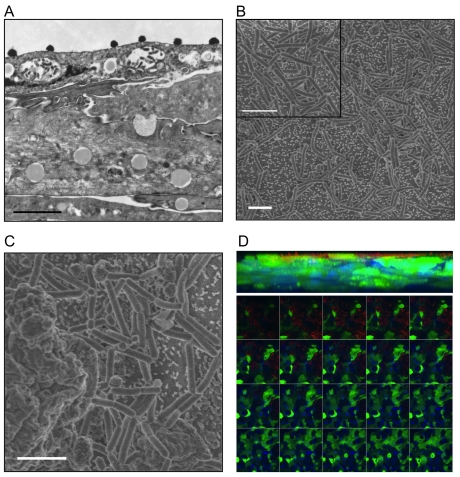
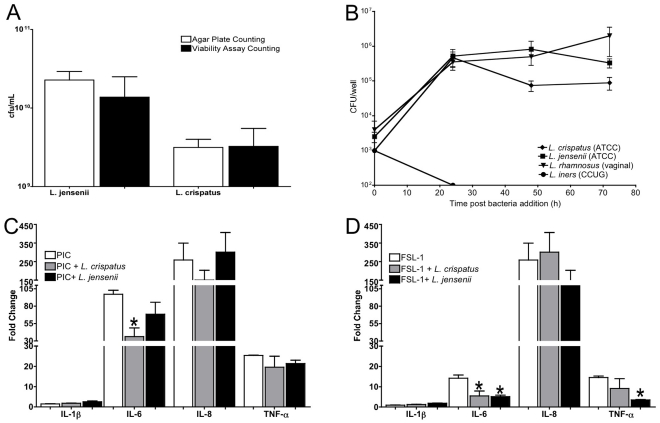
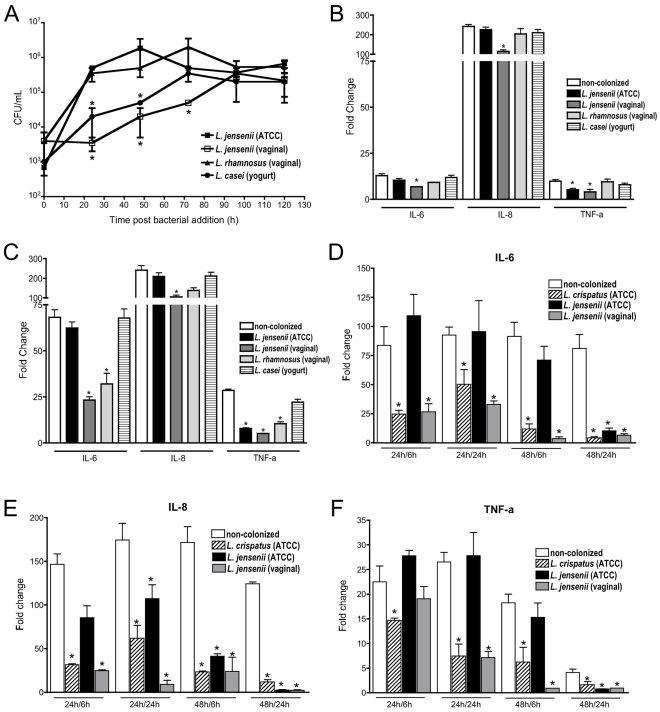
The human vaginal microbiome plays a critical but poorly defined role in reproductive health. Vaginal microbiome alterations are associated with increased susceptibility to sexually-transmitted infections (STI) possibly due to related changes in innate defense responses from epithelial cells. Study of the impact of commensal bacteria on the vaginal mucosal surface has been hindered by current vaginal epithelial cell (VEC) culture systems that lack an appropriate interface between the apical surface of stratified squamous epithelium and the air-filled vaginal lumen. Therefore we developed a reproducible multilayer VEC culture system with an apical (luminal) air-interface that supported colonization with selected commensal bacteria. Multilayer VEC developed tight-junctions and other hallmarks of the vaginal mucosa including predictable proinflammatory cytokine secretion following TLR stimulation. Colonization of multilayers by common vaginal commensals including Lactobacillus crispatus, L. jensenii, and L. rhamnosus led to intimate associations with the VEC exclusively on the apical surface. Vaginal commensals did not trigger cytokine secretion but Staphylococcus epidermidis, a skin commensal, was inflammatory. Lactobacilli reduced cytokine secretion in an isolate-specific fashion following TLR stimulation. This tempering of inflammation offers a potential explanation for increased susceptibility to STI in the absence of common commensals and has implications for testing of potential STI preventatives.
Conflict of interest statement
Figures
References
-
- Eschenbach DA, Thwin SS, Patton DL, Hooton TM, Stapleton AE, et al. Influence of the normal menstrual cycle on vaginal tissue, discharge, and microflora. Clin Infect Dis. 2000;30:901–907. - PubMed
-
- Farage M, Maibach H. Lifetime changes in the vulva and vagina. Arch Gynecol Obstet. 2006;273:195–202. - PubMed
-
- Farage MA, Maibach HI. Morphology and physiological changes of genital skin and mucosa. Curr Probl Dermatol. 2011;40:9–19. - PubMed
-
- Ayehunie S, Cannon C, Lamore S, Kubilus J, Anderson DJ, et al. Organotypic human vaginal-ectocervical tissue model for irritation studies of spermicides, microbicides, and feminine-care products. Toxicol In Vitro. 2006;20:689–698. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
