

Expression patterns of genes involved in sugar metabolism and accumulation during apple fruit development
- PMID: 22412983
- PMCID: PMC3296772
- DOI: 10.1371/journal.pone.0033055
Expression patterns of genes involved in sugar metabolism and accumulation during apple fruit development
Abstract
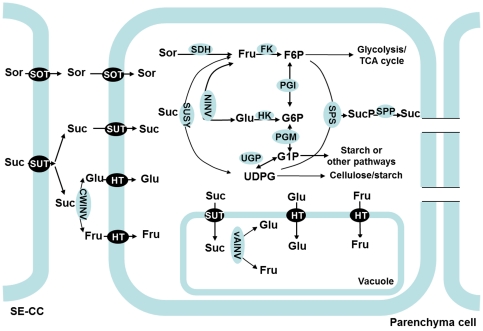
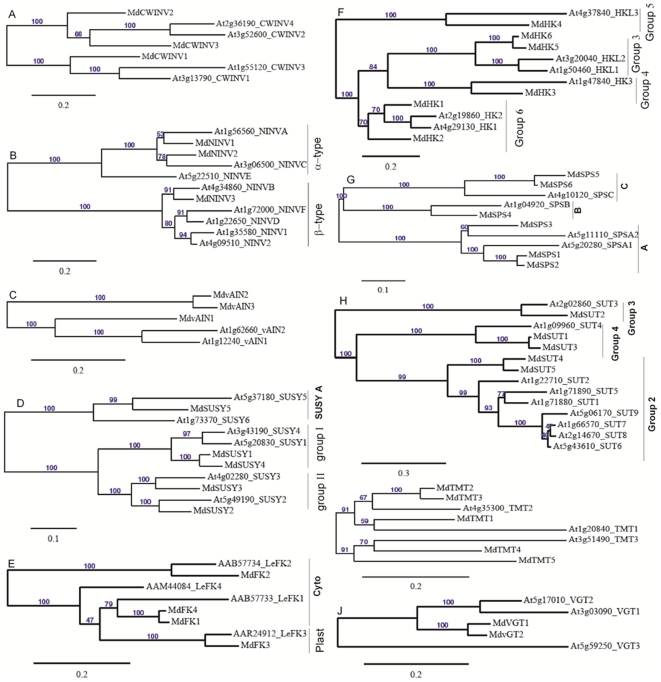
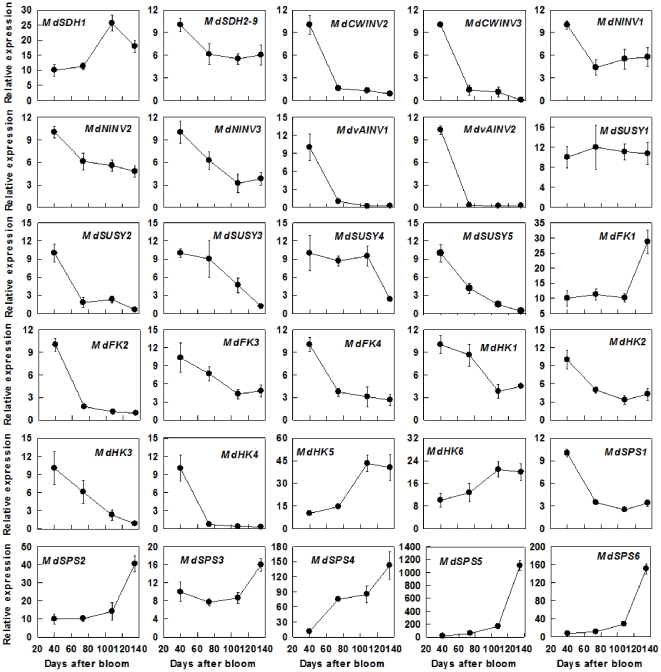
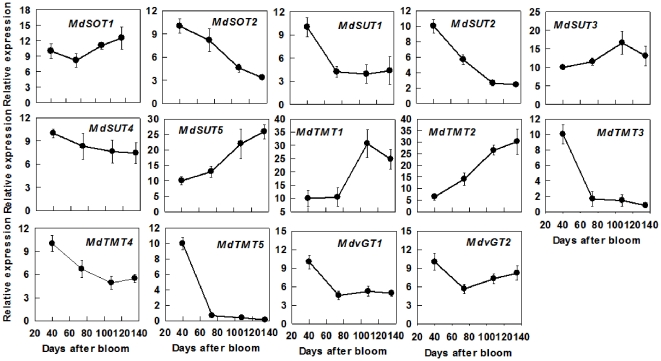
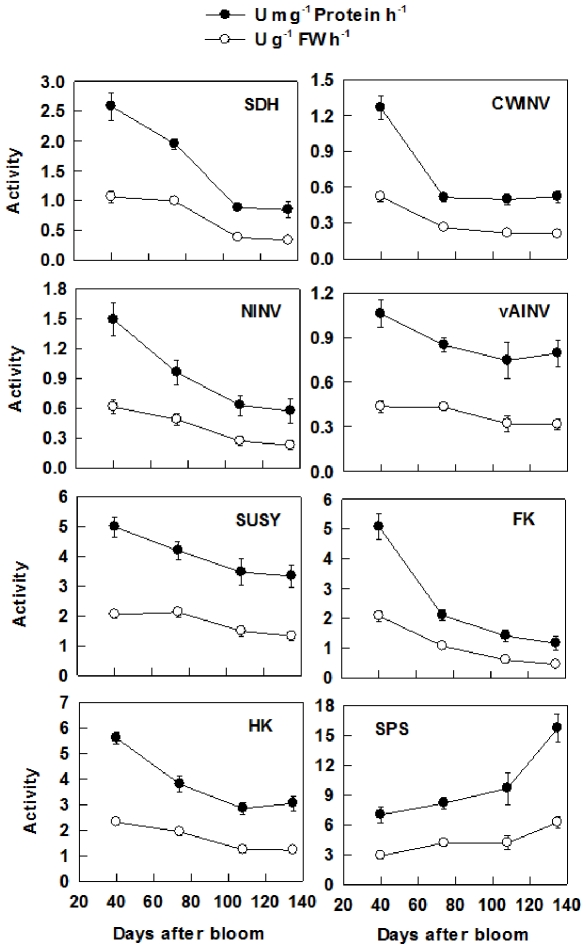
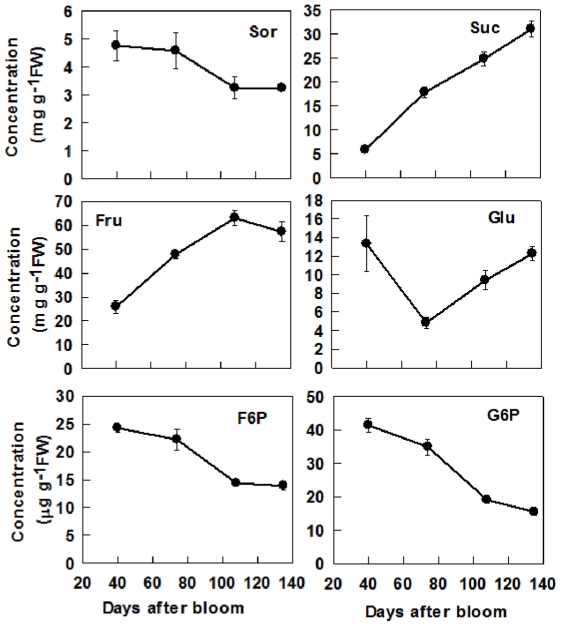
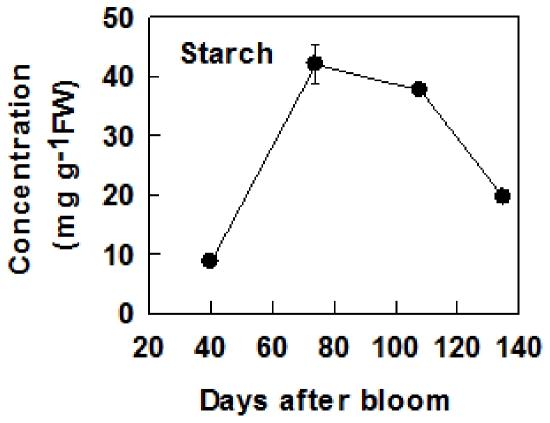
Both sorbitol and sucrose are imported into apple fruit from leaves. The metabolism of sorbitol and sucrose fuels fruit growth and development, and accumulation of sugars in fruit is central to the edible quality of apple. However, our understanding of the mechanisms controlling sugar metabolism and accumulation in apple remains quite limited. We identified members of various gene families encoding key enzymes or transporters involved in sugar metabolism and accumulation in apple fruit using homology searches and comparison of their expression patterns in different tissues, and analyzed the relationship of their transcripts with enzyme activities and sugar accumulation during fruit development. At the early stage of fruit development, the transcript levels of sorbitol dehydrogenase, cell wall invertase, neutral invertase, sucrose synthase, fructokinase and hexokinase are high, and the resulting high enzyme activities are responsible for the rapid utilization of the imported sorbitol and sucrose for fruit growth, with low levels of sugar accumulation. As the fruit continues to grow due to cell expansion, the transcript levels and activities of these enzymes are down-regulated, with concomitant accumulation of fructose and elevated transcript levels of tonoplast monosaccharide transporters (TMTs), MdTMT1 and MdTMT2; the excess carbon is converted into starch. At the late stage of fruit development, sucrose accumulation is enhanced, consistent with the elevated expression of sucrose-phosphate synthase (SPS), MdSPS5 and MdSPS6, and an increase in its total activity. Our data indicate that sugar metabolism and accumulation in apple fruit is developmentally regulated. This represents a comprehensive analysis of the genes involved in sugar metabolism and accumulation in apple, which will serve as a platform for further studies on the functions of these genes and subsequent manipulation of sugar metabolism and fruit quality traits related to carbohydrates.
Conflict of interest statement
Figures
References
-
- Rolland F, Baena-Gonzalez E, Sheen J. Sugar sensing and signaling in plants: conserved and novel mechanisms. Annu Rev Plant Biol. 2006;57:675–709. - PubMed
-
- Ruan Y-L, Jin Y, Yang Y-J, Li G-J, Boyer JS. Sugar input, metabolism, and signaling mediated by invertase: roles in development, yield potential, and response to drought and heat. Mol Plant. 2010;3:942–955. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
