

Growth hormone-induced JAK2 signaling and GH receptor down-regulation: role of GH receptor intracellular domain tyrosine residues
- PMID: 22416081
- PMCID: PMC3339656
- DOI: 10.1210/en.2011-1452
Growth hormone-induced JAK2 signaling and GH receptor down-regulation: role of GH receptor intracellular domain tyrosine residues
Abstract
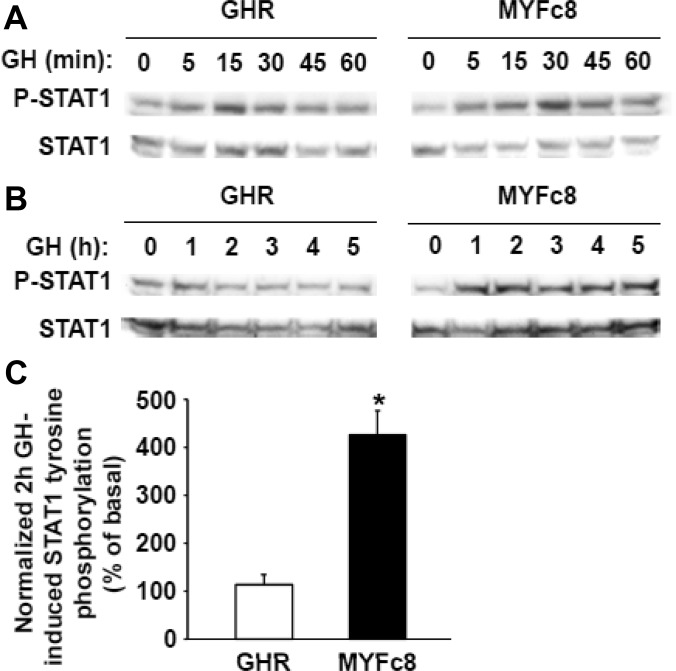
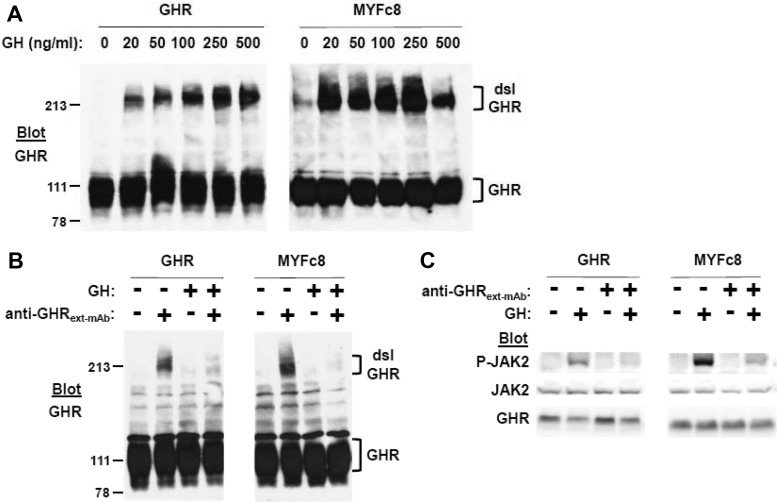
GH receptor (GHR) mediates important somatogenic and metabolic effects of GH. A thorough understanding of GH action requires intimate knowledge of GHR activation mechanisms, as well as determinants of GH-induced receptor down-regulation. We previously demonstrated that a GHR mutant in which all intracellular tyrosine residues were changed to phenylalanine was defective in its ability to activate signal transducer and activator of transcription (STAT)5 and deficient in GH-induced down-regulation, but able to allow GH-induced Janus family of tyrosine kinase 2 (JAK2) activation. We now further characterize the signaling and trafficking characteristics of this receptor mutant. We find that the mutant receptor's extracellular domain conformation and its interaction with GH are indistinguishable from the wild-type receptor. Yet the mutant differs greatly from the wild-type in that GH-induced JAK2 activation is augmented and far more persistent in cells bearing the mutant receptor. Notably, unlike STAT5 tyrosine phosphorylation, GH-induced STAT1 tyrosine phosphorylation is retained and augmented in mutant GHR-expressing cells. The defective receptor down-regulation and persistent JAK2 activation of the mutant receptor do not depend on the sustained presence of GH or on the cell's ability to carry out new protein synthesis. Mutant receptors that exhibit resistance to GH-induced down-regulation are enriched in the disulfide-linked form of the receptor, which reflects the receptor's activated conformation. Furthermore, acute GH-induced internalization, a proximal step in down-regulation, is markedly impaired in the mutant receptor compared to the wild-type receptor. These findings are discussed in the context of determinants and mechanisms of regulation of GHR down-regulation.
Figures




References
-
- Kaplan S. 1999. Hormonal regulation of growth and metabolic effects of growth hormone. In: Kostyo JL, Goodman HM, eds. Handbook of physiology. Chap 5 New York: Oxford University Press; 129–143
-
- Møller N, Jørgensen JO. 2009. Effects of growth hormone on glucose, lipid, and protein metabolism in human subjects. Endocr Rev 30:152–177 - PubMed
-
- Carter-Su C, Schwartz J, Smit LS. 1996. Molecular mechanism of growth hormone action. Annu Rev Physiol 58:187–207 - PubMed
-
- Frank SJ, Gilliland G, Kraft AS, Arnold CS. 1994. Interaction of the growth hormone receptor cytoplasmic domain with the JAK2 tyrosine kinase. Endocrinology 135:2228–2239 - PubMed
-
- Frank SJ, Yi W, Zhao Y, Goldsmith JF, Gilliland G, Jiang J, Sakai I, Kraft AS. 1995. Regions of the JAK2 tyrosine kinase required for coupling to the growth hormone receptor. J Biol Chem 270:14776–14785 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
