

Review
doi: 10.1017/s0952523811000344.
The tasks of amacrine cells
Affiliations
- PMID: 22416289
- PMCID: PMC3652807
- DOI: 10.1017/s0952523811000344
Item in Clipboard
Review
The tasks of amacrine cells
Vis Neurosci.
2012 Jan.
Abstract
Their unique patterns of size, numbers, and stratification indicate that amacrine cells have diverse functions. These are mostly unknown, as studies using imaging and electrophysiological methods have only recently begun. However, some of the events that occur within the amacrine cell population--and some important unresolved puzzles--can be stated purely from structural reasoning.
Copyright © Cambridge University Press, 2012
Figures
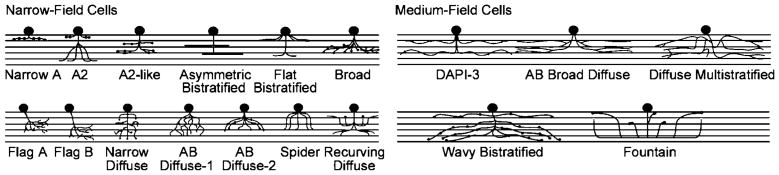
Narrow- and medium-field amacrine cells. The icons represent the levels of stratification of the different amacrine cell types within the IPL and roughly show their spread. Details of the dendrites are represented only when they are a particularly striking feature of the type. For a sense of scale, note that the dendritic field diameters of all of the medium-field cells are near 175 μm. Adapted from Masland (2001).
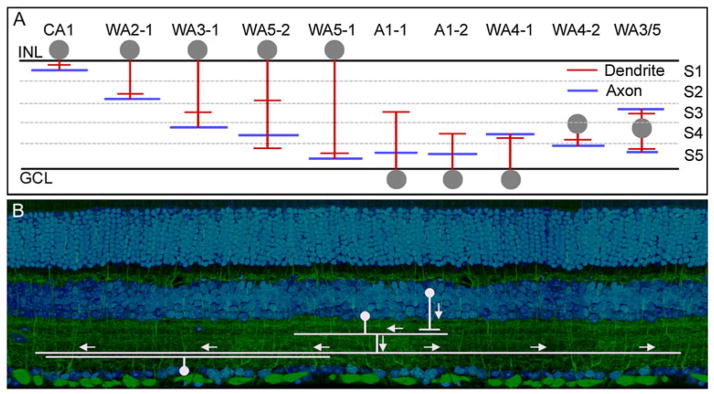
(A) Polyaxonal amacrine cells of the mouse retina. Dendrites are represented in red and axons in blue. Note that these cells are present in most layers of the IPL, and that some have axons and dendrites in different layers. It is not known whether this is an exhaustive list or whether more polyaxonal cells remain to be discovered. These cells were first well described in the retina of the rabbit (Vaney et al., 1988; Famiglietti, 1992a,b,c). Lin and Masland (2006) surveyed GFP-M retinas (Feng et al., 2000) for the presence of long-range amacrine cells. Badea and Nathans (2004) and Gustincich et al. (1997) used different genetically based strategies and their results are included here. No standard nomenclature exists; for details, see the original publications. The lateral spread of the processes is not represented, except to indicate that the spread of the dendritic arbor in these cells is narrower than the spread of the axonal arbor. (B) Polyaxonal cells transmit information from one or a few types of bipolar and amacrine cells to specific other groups of cells, here illustrated for one type of bipolar cell and one type of ganglion cell. This is shown by both the laminar structure of the polyaxonal cells, and the fact that their physiological effects are restricted to subsets of ganglion cells (Olveczky et al., 2003; Roska & Werblin, 2003).

The relative sizes of amacrine cells and ganglion cells. This is a whole-mount view, with the boundaries of the dendritic fields outlined. A ganglion cell of roughly average dendritic field diameter (250 μm) for the rabbit is shown by the large dark outline. A mosaic of narrow-field (left) or wide-field (right) amacrine cells is shown in white. Many narrow-field amacrine cells are present within the dendritic field of this ganglion cell; they must create some sort of a functional subunit within the receptive field, but this functionality has been little explored. In contrast, medium-field amacrine cells have a confusing overlap with the dendritic field of the ganglion cell; it is not obvious how the activities of these two cells can be spatially integrated.
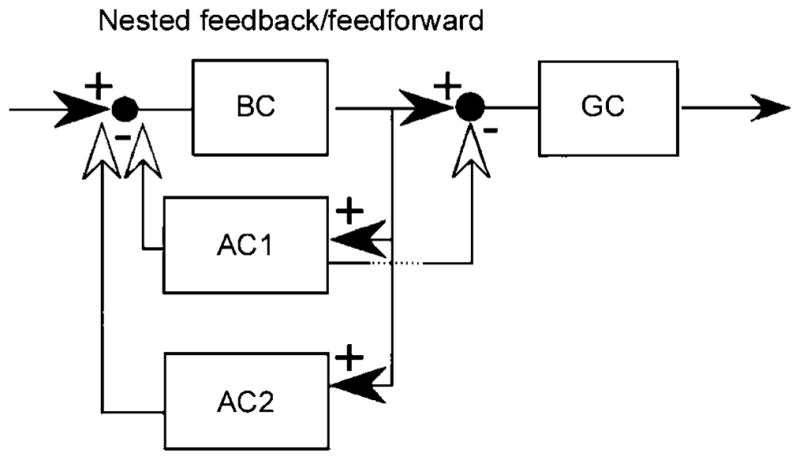
One of the many possible consequences of synapses among bipolar, amacrine, and ganglion cells. The one shown here was termed “nested feedforward inhibition” by Marc and Liu (2000), who pointed out that several such logics are possible. Reproduced by permission from the Journal of Comparative Neurology.
References
-
- Badea TC, Nathans J. Quantitative analysis of neuronal morphologies in the mouse retina visualized by using a genetically directed reporter. The Journal of Comparative Neurology. 2004;480:331–351. - PubMed
-
- Briggman KL, Denk W. Towards neural circuit reconstruction with volume electron microscopy techniques. Current Opinion in Neurobiology. 2006;16:562–570. - PubMed
-
- Burnside B. Light and circadian regulation of retinomotor movement. Progress in Brain Research. 2001;131:477–485. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
