

Host cell deformability is linked to transmission in the human malaria parasite Plasmodium falciparum
- PMID: 22417683
- PMCID: PMC3376226
- DOI: 10.1111/j.1462-5822.2012.01786.x
Host cell deformability is linked to transmission in the human malaria parasite Plasmodium falciparum
Abstract
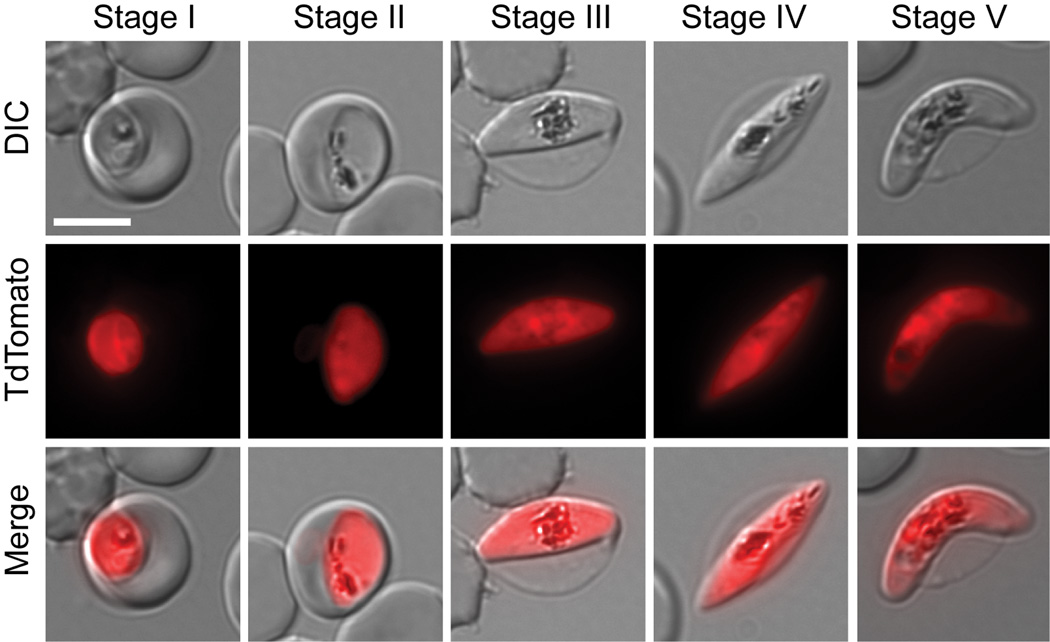
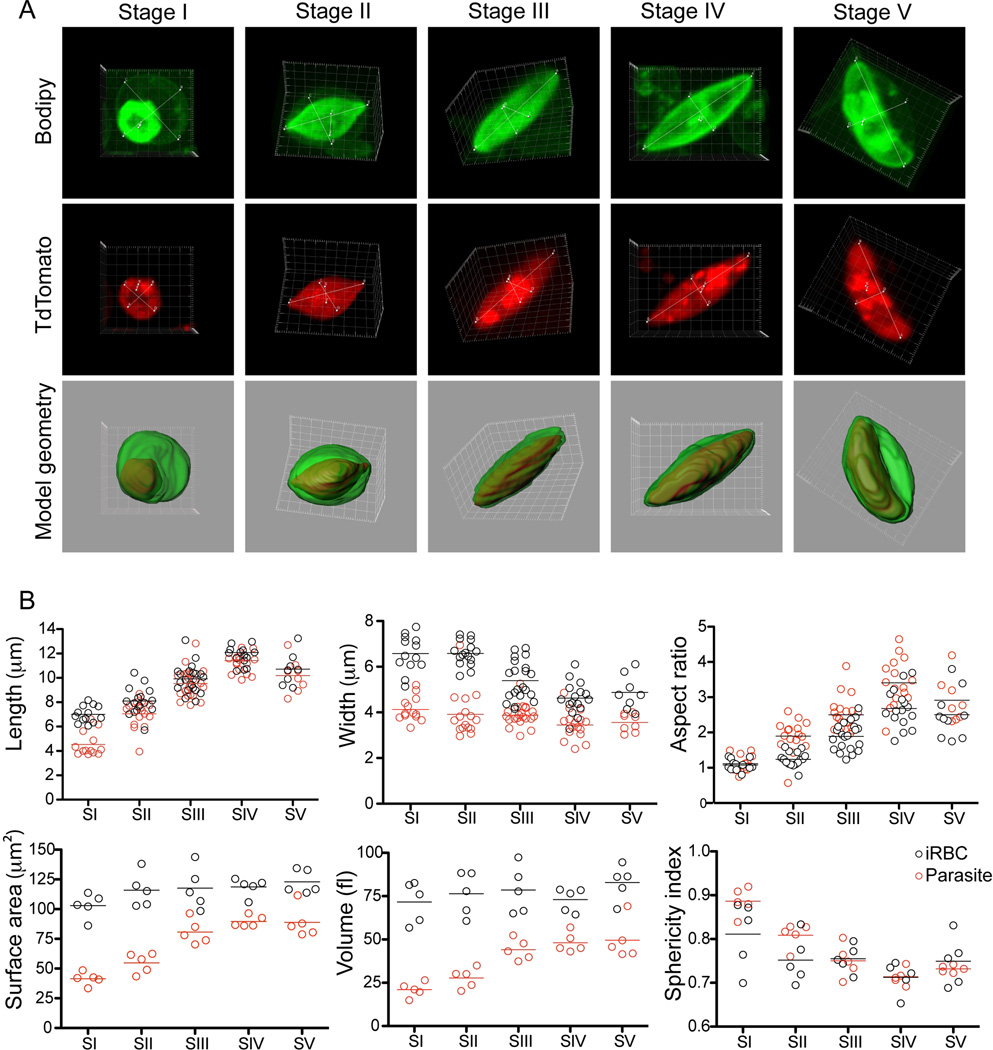
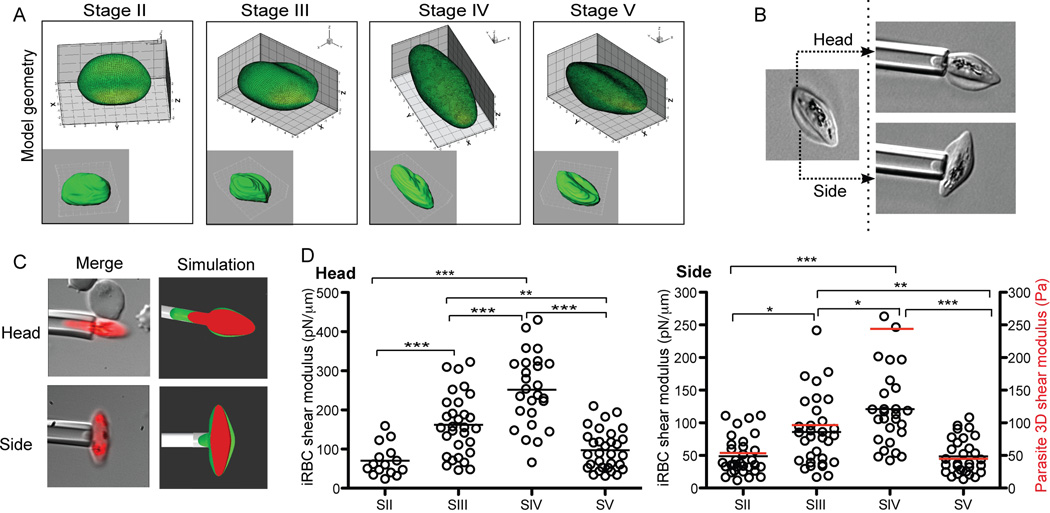
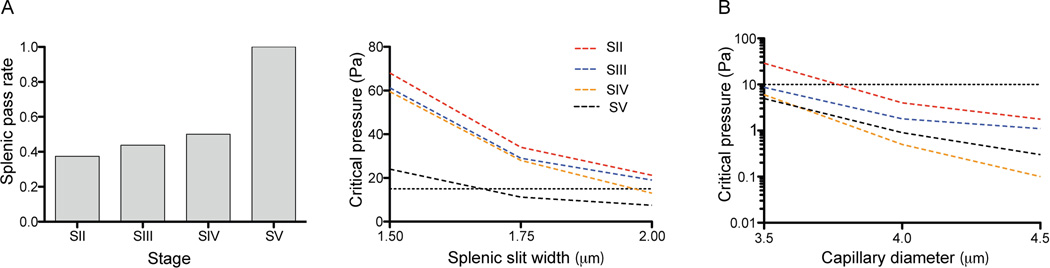
Gametocyte maturation in Plasmodium falciparum is a critical step in the transmission of malaria. While the majority of parasites proliferate asexually in red blood cells, a small fraction of parasites undergo sexual conversion and mature over 2 weeks to become competent for transmission to a mosquito vector. Immature gametocytes sequester in deep tissues while mature stages must be able to circulate, pass the spleen and present themselves to the mosquito vector in order to complete transmission. Sequestration of asexual red blood cell stage parasites has been investigated in great detail. These studies have demonstrated that induction of cytoadherence properties through specific receptor-ligand interactions coincides with a significant increase in host cell stiffness. In contrast, the adherence and biophysical properties of gametocyte-infected red blood cells have not been studied systematically. Utilizing a transgenic line for 3D live imaging, in vitro capillary assays and 3D finite element whole cell modelling, we studied the role of cellular deformability in determining the circulatory characteristics of gametocytes. Our analysis shows that the red blood cell deformability of immature gametocytes displays an overall decrease followed by rapid restoration in mature gametocytes. Intriguingly, simulations suggest that along with deformability variations, the morphological changes of the parasite may play an important role in tissue distribution in vivo. Taken together, we present a model, which suggests that mature but not immature gametocytes circulate in the peripheral blood for uptake in the mosquito blood meal and transmission to another human host thus ensuring long-term survival of the parasite.
© 2012 Blackwell Publishing Ltd.
Figures
References
-
- Cokelet GR. Dynamics of erythrocyte motion in filtration tests and in vivo flow. Scand J Clin Lab Invest Suppl. 1981;156:77–82. - PubMed
-
- Dao M, Lim CT, Suresh S. Mechanics of the human red blood cell deformed by optical tweezers. Journal of the Mechanics and Physics of Solids. 2003;51:2259–2280.
-
- David PH, Hommel M, Miller LH, Udeinya IJ, Oligino LD. Parasite sequestration in Plasmodium falciparum malaria: spleen and antibody modulation of cytoadherence of infected erythrocytes. Proceedings of the National Academy of Sciences of the United States of America. 1983;80:5075–5079. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
