

Hepatocyte-stellate cell cross-talk in the liver engenders a permissive inflammatory microenvironment that drives progression in hepatocellular carcinoma
- PMID: 22419664
- PMCID: PMC3498759
- DOI: 10.1158/0008-5472.CAN-11-3317
Hepatocyte-stellate cell cross-talk in the liver engenders a permissive inflammatory microenvironment that drives progression in hepatocellular carcinoma
Abstract
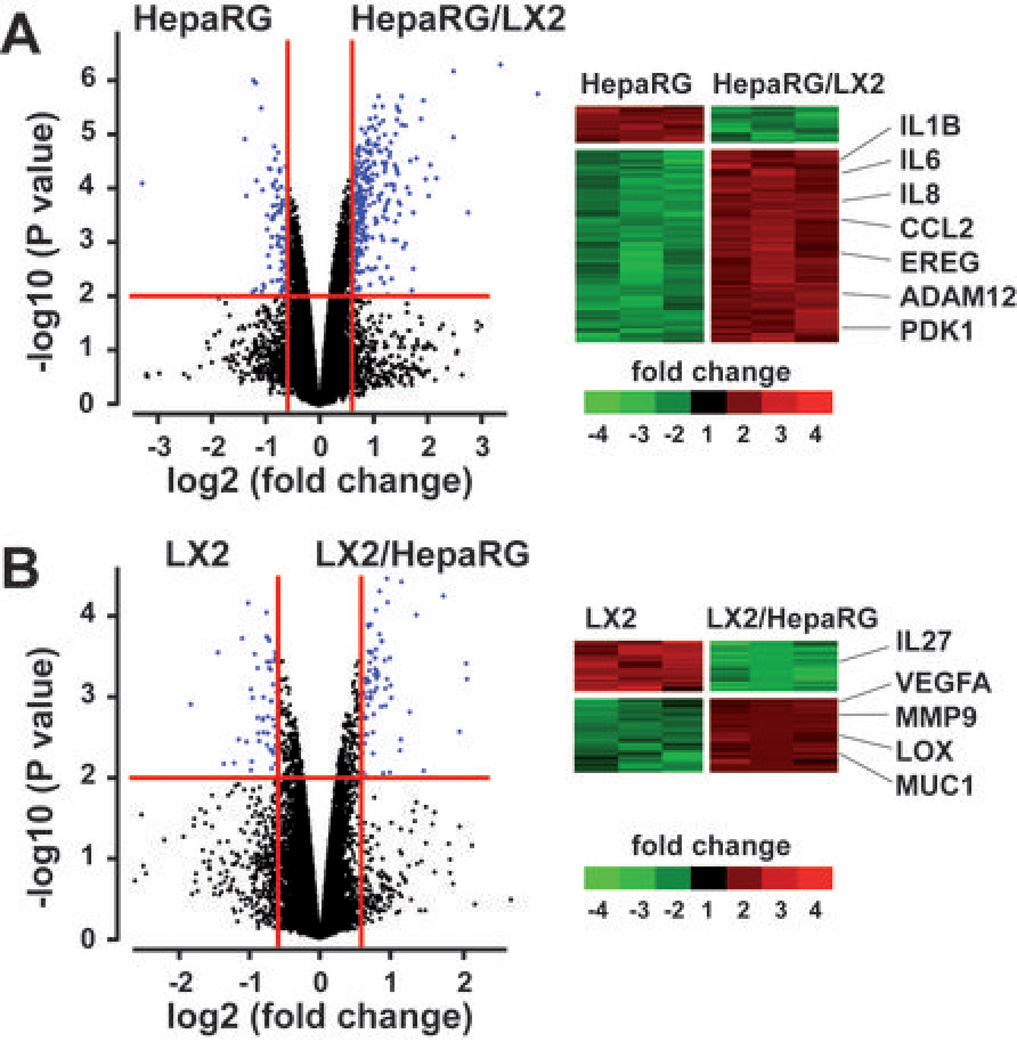
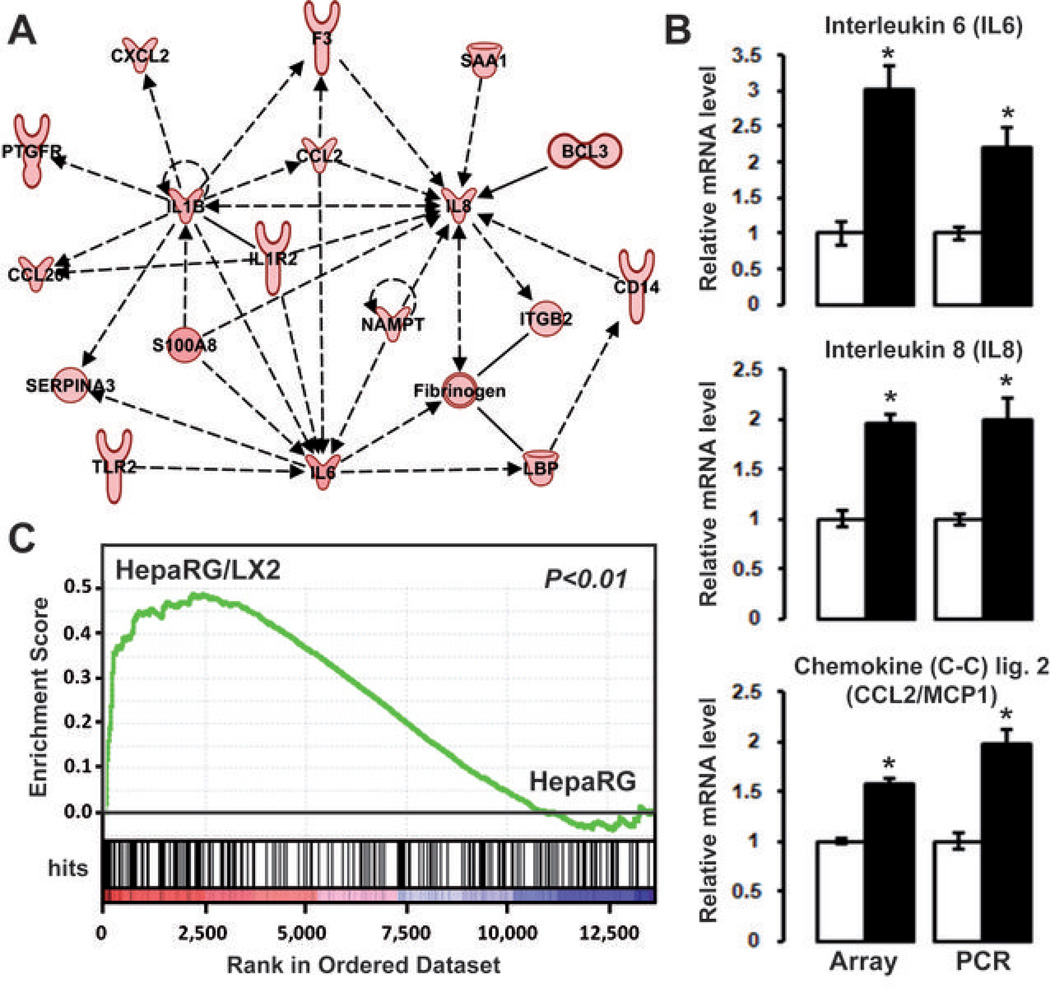
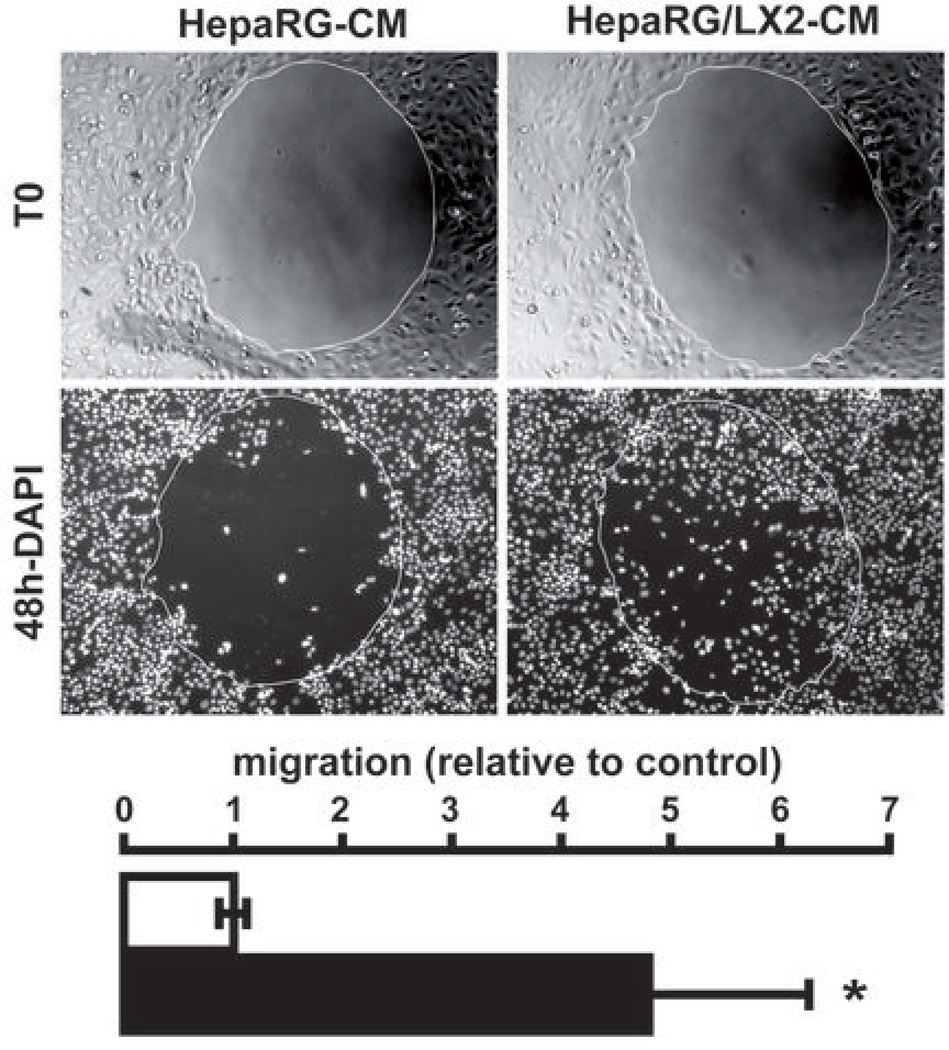
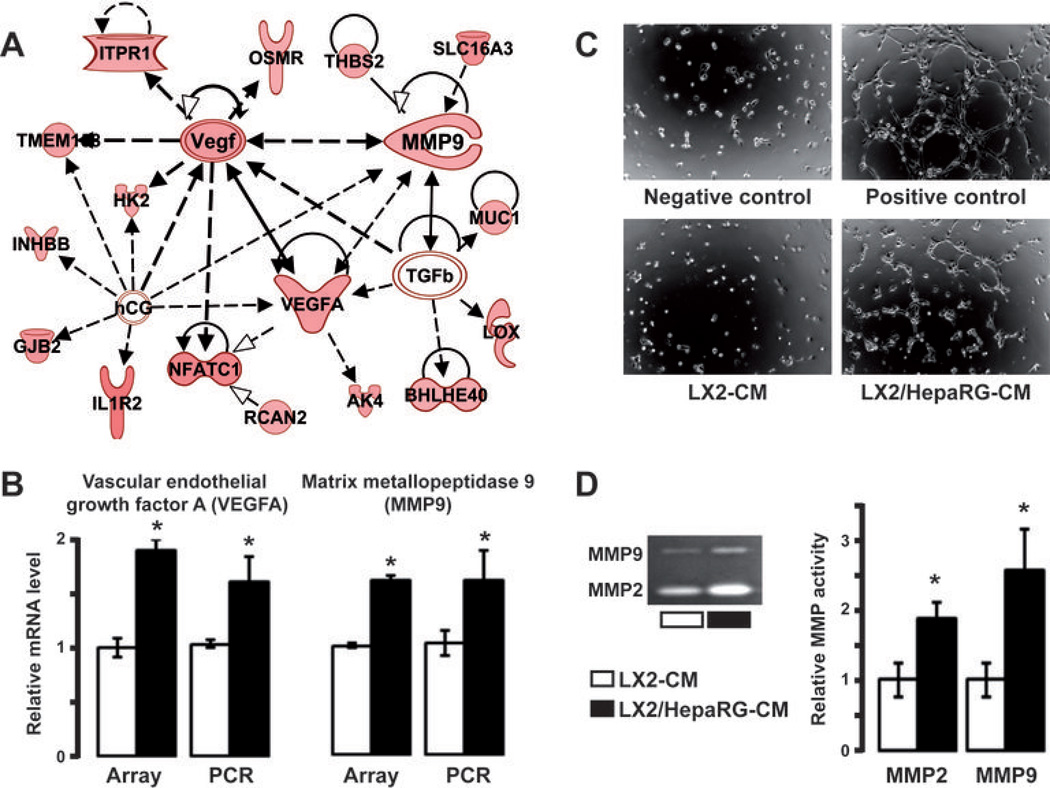
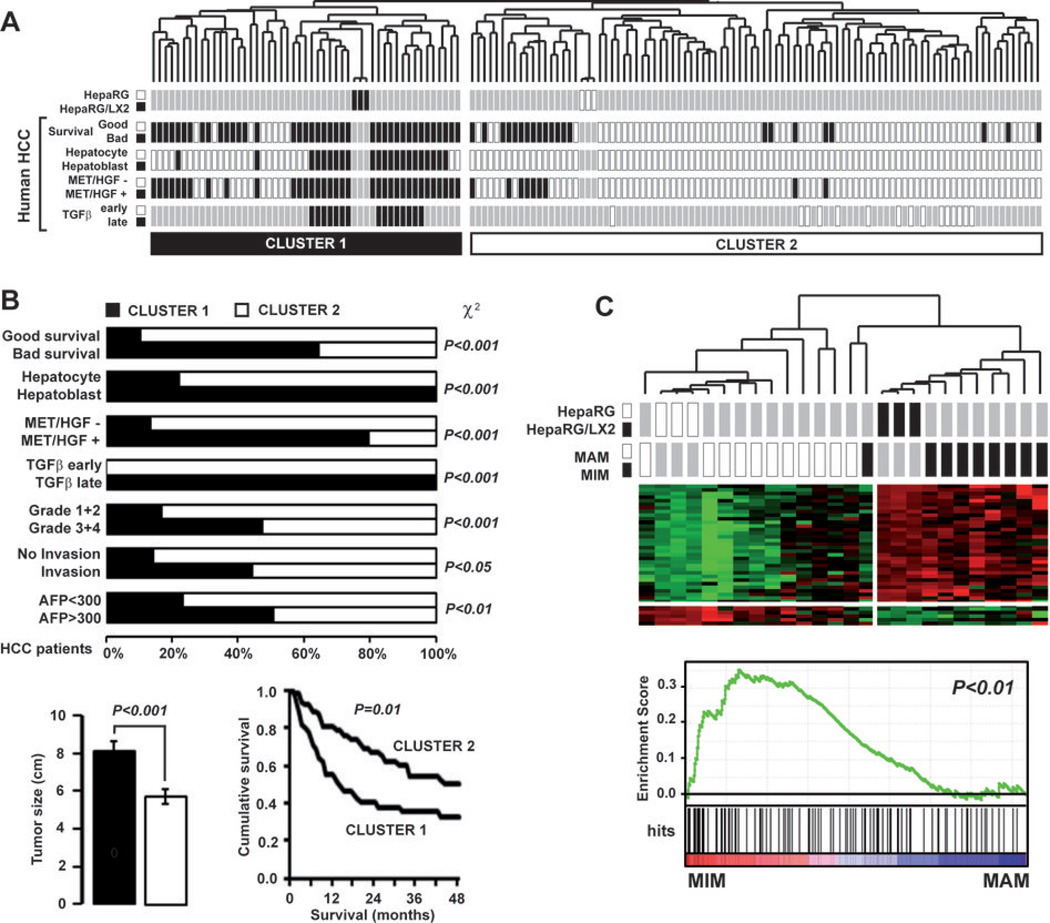
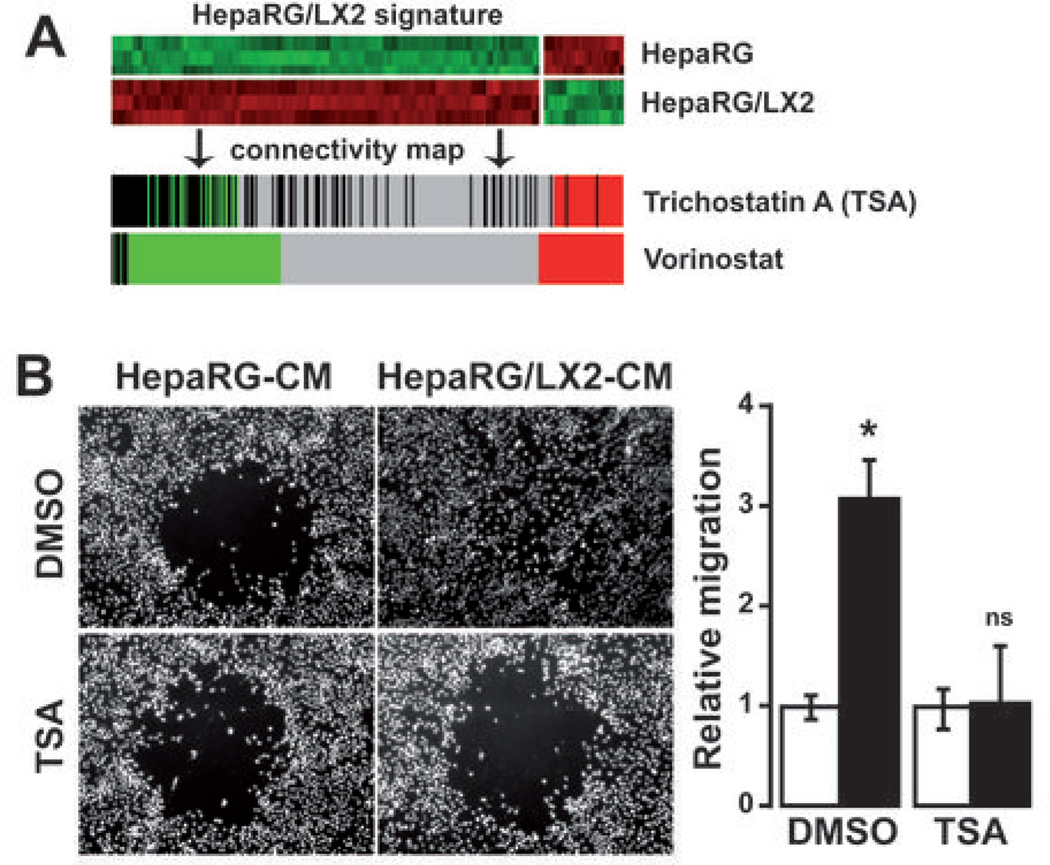
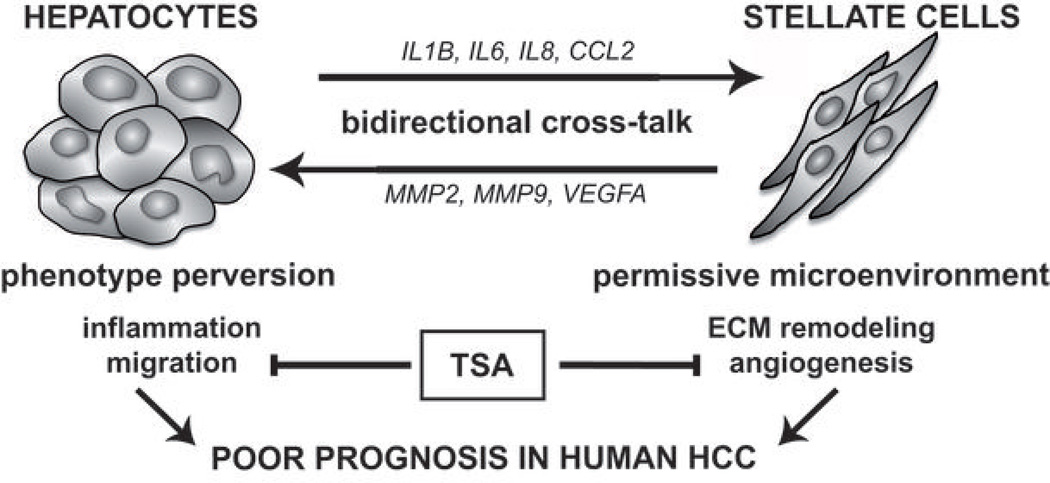
Many solid malignant tumors arise on a background of inflamed and/or fibrotic tissues, features that are found in more than 80% hepatocellular carcinomas (HCC). Activated hepatic stellate cells (HSC) play a critical role in fibrogenesis associated with HCC onset and progression, yet their functional impact on hepatocyte fate remains largely unexplored. Here, we used a coculture model to investigate the cross-talk between hepatocytes (human hepatoma cells) and activated human HSCs. Unsupervised genome-wide expression profiling showed that hepatocyte-HSC cross-talk is bidirectional and results in the deregulation of functionally relevant gene networks. Notably, coculturing increased the expression of proinflammatory cytokines and modified the phenotype of hepatocytes toward motile cells. Hepatocyte-HSC cross-talk also generated a permissive proangiogenic microenvironment, particularly by inducing VEGFA and matrix metalloproteinase (MMP)9 expression in HSCs. An integrative genomic analysis revealed that the expression of genes associated with hepatocyte-HSC cross-talk correlated with HCC progression in mice and was predictive of a poor prognosis and metastasis propensity in human HCCs. Interestingly, the effects of cross-talk on migration and angiogenesis were reversed by the histone deacetylase inhibitor trichostatin A. Our findings, therefore, indicate that the cross-talk between hepatoma cells and activated HSCs is an important feature of HCC progression, which may be targeted by epigenetic modulation.
©2012 AACR.
Figures
References
-
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–674. - PubMed
-
- Bhowmick NA, Chytil A, Plieth D, et al. TGF-beta signaling in fibroblasts modulates the oncogenic potential of adjacent epithelia. Science. 2004;303:848–851. - PubMed
-
- Polyak K, Haviv I, Campbell IG. Co-evolution of tumor cells and their microenvironment. Trends Genet. 2009;25:30–38. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
