

Generation and physiological roles of linear ubiquitin chains
- PMID: 22420778
- PMCID: PMC3305636
- DOI: 10.1186/1741-7007-10-23
Generation and physiological roles of linear ubiquitin chains
Abstract
Ubiquitination now ranks with phosphorylation as one of the best-studied post-translational modifications of proteins with broad regulatory roles across all of biology. Ubiquitination usually involves the addition of ubiquitin chains to target protein molecules, and these may be of eight different types, seven of which involve the linkage of one of the seven internal lysine (K) residues in one ubiquitin molecule to the carboxy-terminal diglycine of the next. In the eighth, the so-called linear ubiquitin chains, the linkage is between the amino-terminal amino group of methionine on a ubiquitin that is conjugated with a target protein and the carboxy-terminal carboxy group of the incoming ubiquitin. Physiological roles are well established for K48-linked chains, which are essential for signaling proteasomal degradation of proteins, and for K63-linked chains, which play a part in recruitment of DNA repair enzymes, cell signaling and endocytosis. We focus here on linear ubiquitin chains, how they are assembled, and how three different avenues of research have indicated physiological roles for linear ubiquitination in innate and adaptive immunity and suppression of inflammation.
Figures
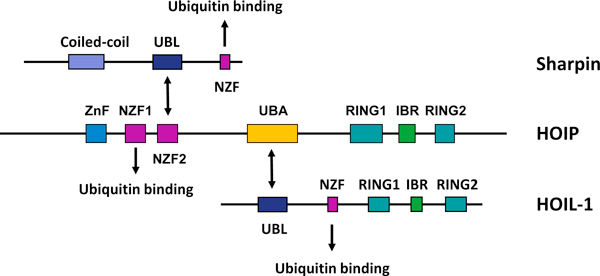
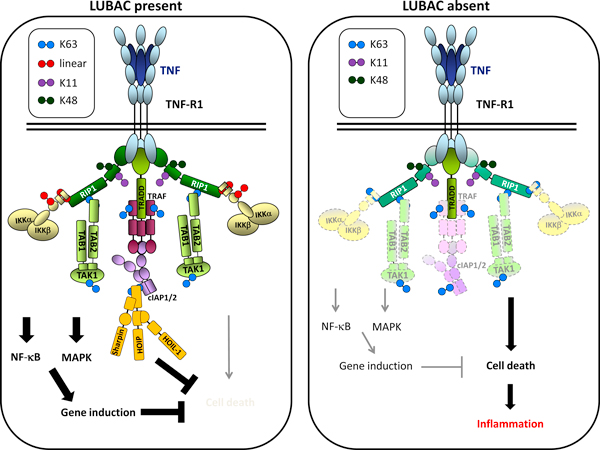
References
-
- Hershko A, Heller H, Elias S, Ciechanover A. Components of ubiquitin-protein ligase system. Resolution, affinity purification, and role in protein breakdown. J Biol Chem. 1983;258:8206–8214. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
