

Ancient deuterostome origins of vertebrate brain signalling centres
- PMID: 22422262
- PMCID: PMC3719855
- DOI: 10.1038/nature10838
Ancient deuterostome origins of vertebrate brain signalling centres
Abstract
Neuroectodermal signalling centres induce and pattern many novel vertebrate brain structures but are absent, or divergent, in invertebrate chordates. This has led to the idea that signalling-centre genetic programs were first assembled in stem vertebrates and potentially drove morphological innovations of the brain. However, this scenario presumes that extant cephalochordates accurately represent ancestral chordate characters, which has not been tested using close chordate outgroups. Here we report that genetic programs homologous to three vertebrate signalling centres-the anterior neural ridge, zona limitans intrathalamica and isthmic organizer-are present in the hemichordate Saccoglossus kowalevskii. Fgf8/17/18 (a single gene homologous to vertebrate Fgf8, Fgf17 and Fgf18), sfrp1/5, hh and wnt1 are expressed in vertebrate-like arrangements in hemichordate ectoderm, and homologous genetic mechanisms regulate ectodermal patterning in both animals. We propose that these genetic programs were components of an unexpectedly complex, ancient genetic regulatory scaffold for deuterostome body patterning that degenerated in amphioxus and ascidians, but was retained to pattern divergent structures in hemichordates and vertebrates.
© 2012 Macmillan Publishers Limited. All rights reserved
Figures
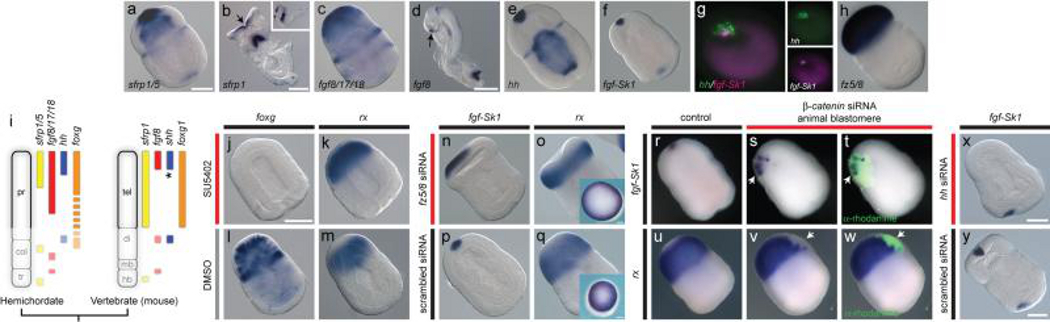
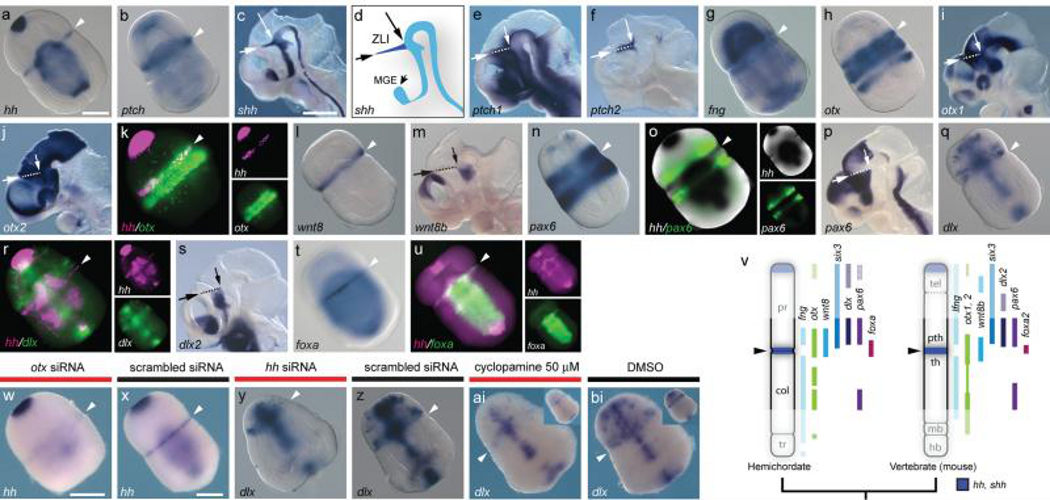
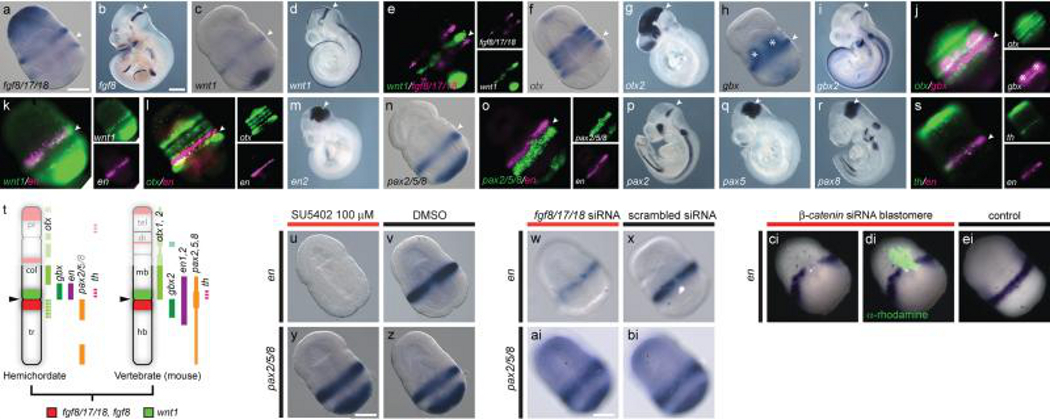
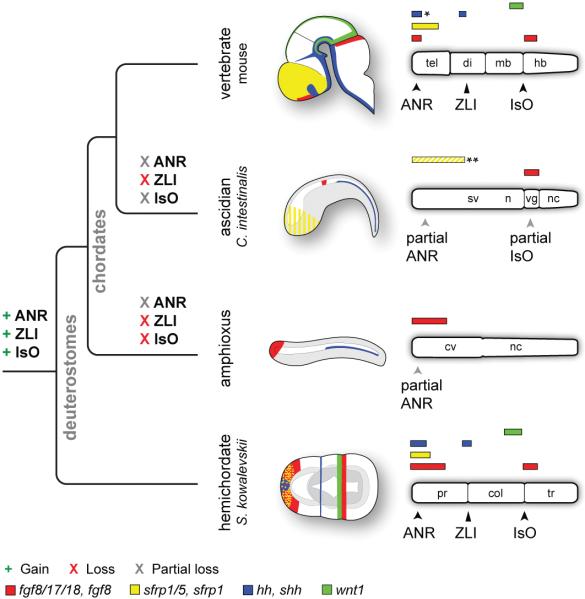
Comment in
-
Developmental biology: a brainy background.Nature. 2012 Mar 14;483(7389):280. doi: 10.1038/483280a. Nature. 2012. PMID: 22422257 No abstract available.
References
-
- Echevarria D, Vieira C, Gimeno L, Martinez S. Neuroepithelial secondary organizers and cell fate specification in the developing brain. Brain Res Brain Res Rev. 2003;43:179–191. doi:S0165017303002078 [pii] - PubMed
-
- Wurst W, Bally-Cuif L. Neural plate patterning: upstream and downstream of the isthmic organizer. Nat Rev Neurosci. 2001;2:99–108. doi:10.1038/35053516. - PubMed
-
- Wicht H, Lacalli TC. The nervous system of amphioxus: structure, development, and evolutionary significance. Canadian Journal of Zoology. 2005;83:122–150. doi:10.1139/z04-163.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
