

Ribosome profiling shows that miR-430 reduces translation before causing mRNA decay in zebrafish
- PMID: 22422859
- PMCID: PMC3547538
- DOI: 10.1126/science.1215704
Ribosome profiling shows that miR-430 reduces translation before causing mRNA decay in zebrafish
Abstract
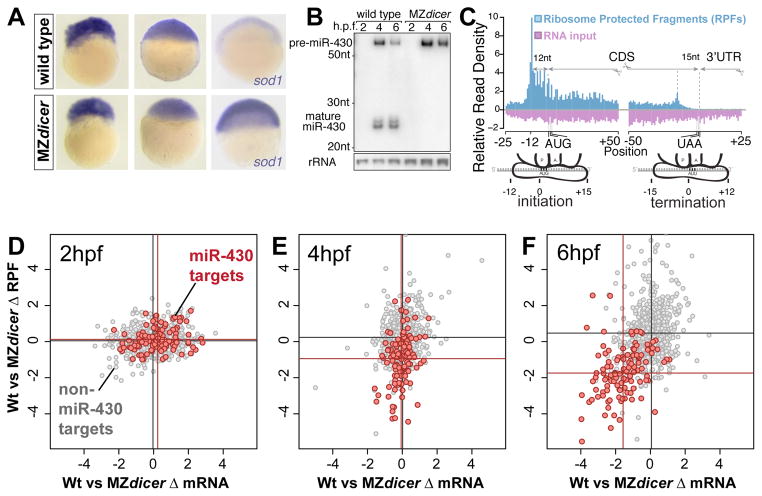
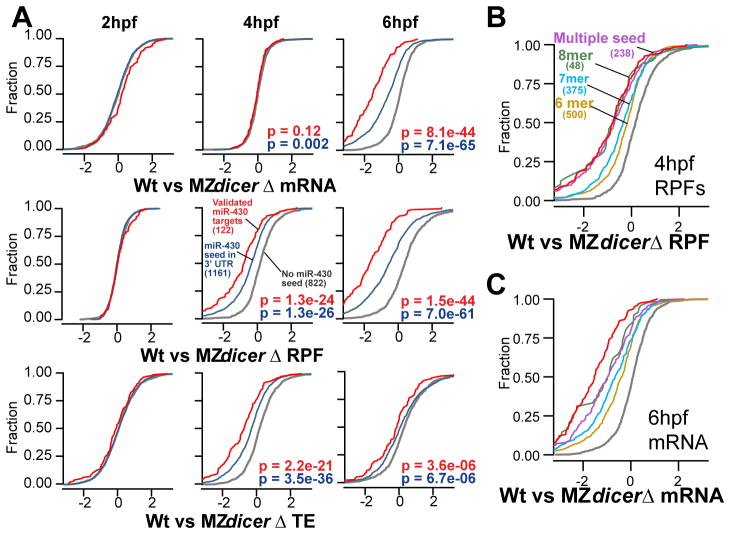
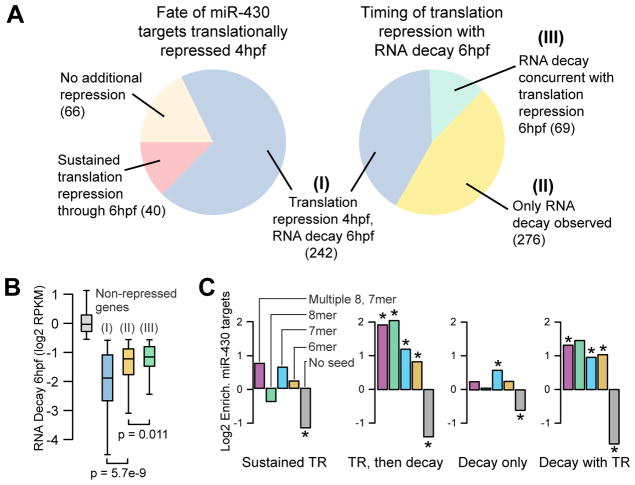
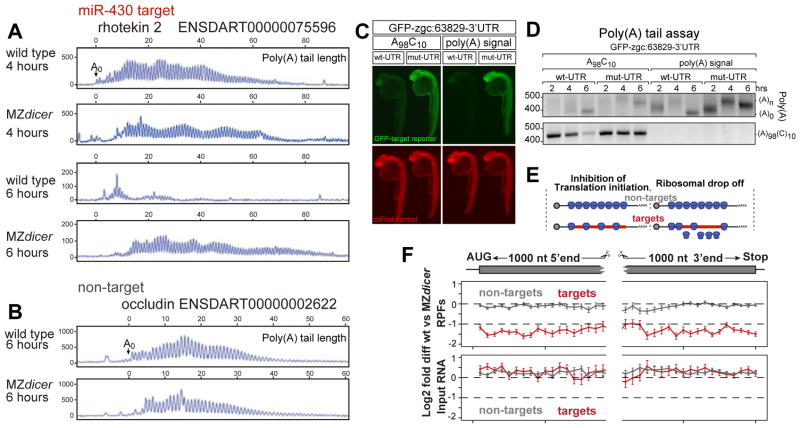
MicroRNAs regulate gene expression through deadenylation, repression, and messenger RNA (mRNA) decay. However, the contribution of each mechanism in non-steady-state situations remains unclear. We monitored the impact of miR-430 on ribosome occupancy of endogenous mRNAs in wild-type and dicer mutant zebrafish embryos and found that miR-430 reduces the number of ribosomes on target mRNAs before causing mRNA decay. Translational repression occurs before complete deadenylation, and disrupting deadenylation with use of an internal polyadenylate tail did not block target repression. Lastly, we observed that ribosome density along the length of the message remains constant, suggesting that translational repression occurs by reducing the rate of initiation rather than affecting elongation or causing ribosomal drop-off. These results show that miR-430 regulates translation initiation before inducing mRNA decay during zebrafish development.
Figures
Comment in
-
Small RNAs: miRNAs' strict schedule.Nat Rev Genet. 2012 May 3;13(6):378. doi: 10.1038/nrg3251. Nat Rev Genet. 2012. PMID: 22552258 No abstract available.
-
miRNAs' strict schedule.Nat Rev Mol Cell Biol. 2012 May 3;13(6):340-1. doi: 10.1038/nrm3354. Nat Rev Mol Cell Biol. 2012. PMID: 22552328 No abstract available.
References
-
- Fabian MR, Sonenberg N, Filipowicz W. Regulation of mRNA translation and stability by microRNAs. Annu Rev Biochem. 2010;79:351. - PubMed
-
- Pillai RS, et al. Inhibition of translational initiation by Let-7 MicroRNA in human cells. Science. 2005;309:1573. - PubMed
-
- Olsen PH, Ambros V. The lin-4 regulatory RNA controls developmental timing in Caenorhabditis elegans by blocking LIN-14 protein synthesis after the initiation of translation. Dev Biol. 1999;216:671. - PubMed
-
- Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75:843. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
