

Autophagy protein Rubicon mediates phagocytic NADPH oxidase activation in response to microbial infection or TLR stimulation
- PMID: 22423966
- PMCID: PMC3616771
- DOI: 10.1016/j.chom.2012.01.018
Autophagy protein Rubicon mediates phagocytic NADPH oxidase activation in response to microbial infection or TLR stimulation
Abstract
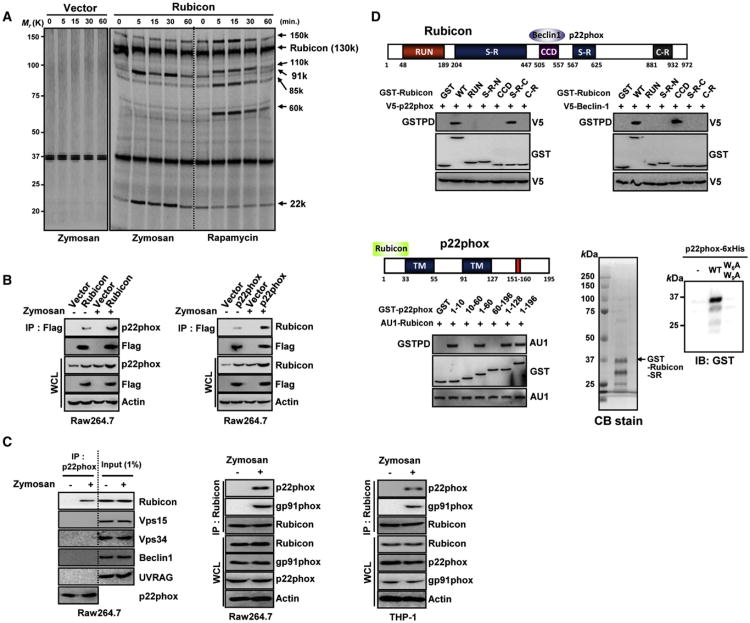
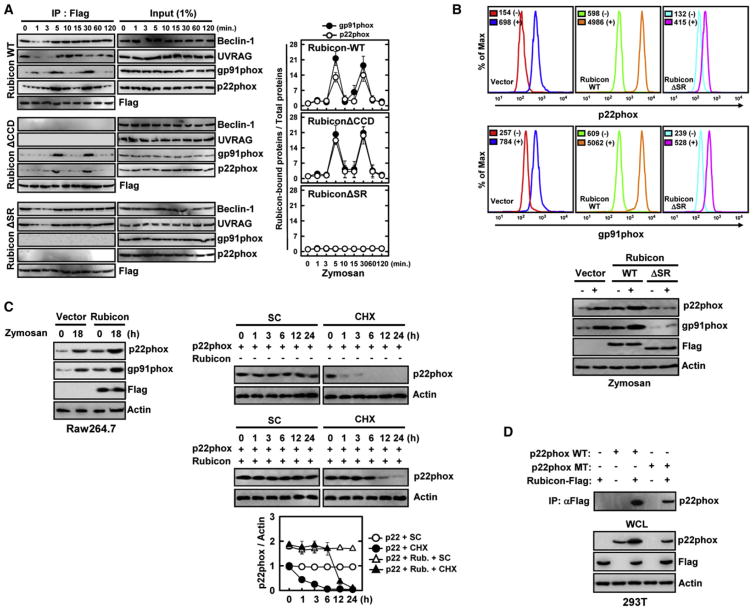
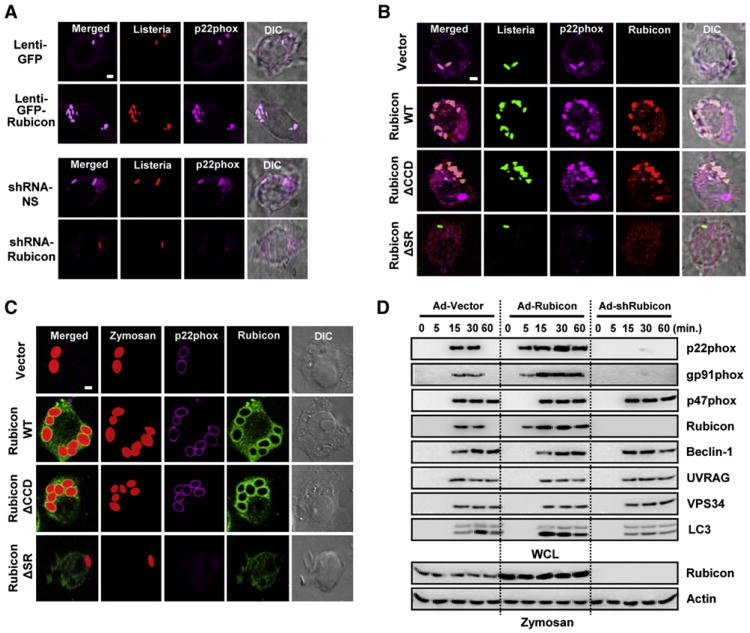
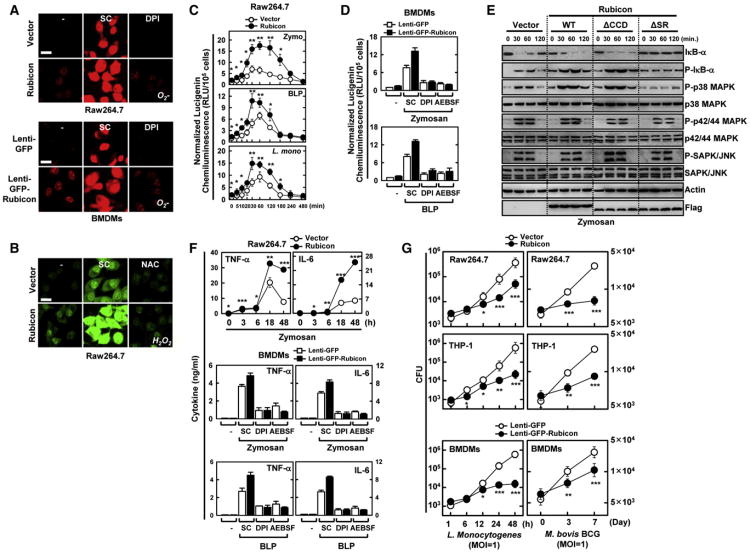
Phagocytosis and autophagy are two important and related arms of the host's first-line defense against microbial invasion. Rubicon is a RUN domain containing cysteine-rich protein that functions as part of a Beclin-1-Vps34-containing autophagy complex. We report that Rubicon is also an essential, positive regulator of the NADPH oxidase complex. Upon microbial infection or Toll-like-receptor 2 (TLR2) activation, Rubicon interacts with the p22phox subunit of the NADPH oxidase complex, facilitating its phagosomal trafficking to induce a burst of reactive oxygen species (ROS) and inflammatory cytokines. Consequently, ectopic expression or depletion of Rubicon profoundly affected ROS, inflammatory cytokine production, and subsequent antimicrobial activity. Rubicon's actions in autophagy and in the NADPH oxidase complex are functionally and genetically separable, indicating that Rubicon functions in two ancient innate immune machineries, autophagy and phagocytosis, depending on the environmental stimulus. Rubicon may thus be pivotal to generating an optimal intracellular immune response against microbial infection.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Crossing the Rubicon: new roads lead to host defense.Cell Host Microbe. 2012 Mar 15;11(3):221-3. doi: 10.1016/j.chom.2012.02.005. Cell Host Microbe. 2012. PMID: 22423961 Free PMC article.
References
-
- Bedard K, Krause KH. The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol Rev. 2007;87:245–313. - PubMed
-
- Deretic V, Klionsky DJ. How cells clean house. Sci Am. 2008;298:74–81. - PubMed
-
- Fang FC. Antimicrobial reactive oxygen and nitrogen species: concepts and controversies. Nat Rev Microbiol. 2004;2:820–832. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01 DE019085/DE/NIDCR NIH HHS/United States
- R01 CA109697/CA/NCI NIH HHS/United States
- R01 CA140964/CA/NCI NIH HHS/United States
- AI073099/AI/NIAID NIH HHS/United States
- AI083025/AI/NIAID NIH HHS/United States
- R01 CA082057/CA/NCI NIH HHS/United States
- AI083841/AI/NIAID NIH HHS/United States
- R01 CA091819/CA/NCI NIH HHS/United States
- U19 AI083025/AI/NIAID NIH HHS/United States
- R01 CA115284/CA/NCI NIH HHS/United States
- R01 AI073099/AI/NIAID NIH HHS/United States
- CA140964/CA/NCI NIH HHS/United States
- R21 AI083841/AI/NIAID NIH HHS/United States
- R01 HL110609/HL/NHLBI NIH HHS/United States
- DE019085/DE/NIDCR NIH HHS/United States
- CA31363/CA/NCI NIH HHS/United States
- HL110609/HL/NHLBI NIH HHS/United States
- R01 CA106156/CA/NCI NIH HHS/United States
- CA082057/CA/NCI NIH HHS/United States
- CA115284/CA/NCI NIH HHS/United States
- R01 CA031363/CA/NCI NIH HHS/United States
- R01 CA086841/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
