

Natural product biosynthetic gene diversity in geographically distinct soil microbiomes
- PMID: 22427492
- PMCID: PMC3346347
- DOI: 10.1128/AEM.00102-12
Natural product biosynthetic gene diversity in geographically distinct soil microbiomes
Abstract
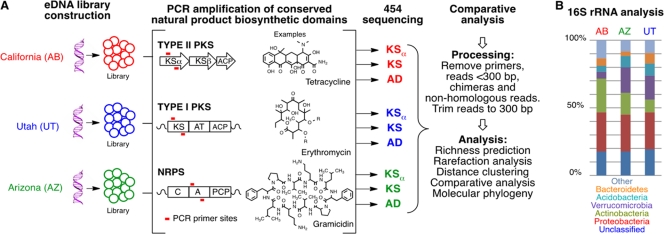
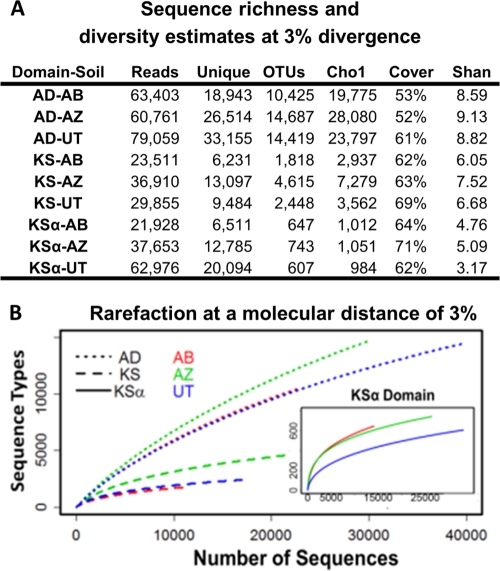
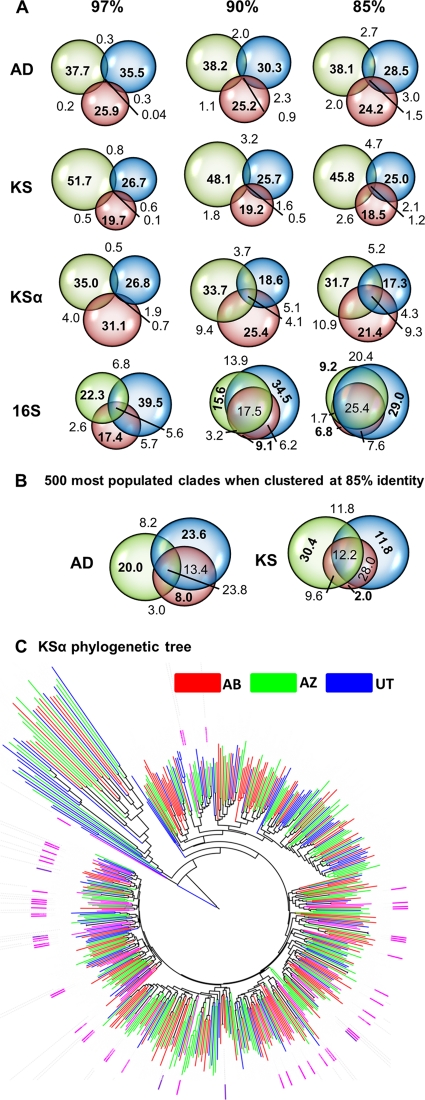
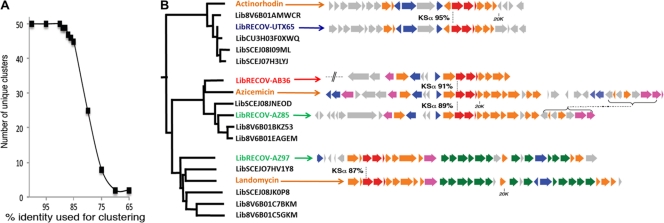
The number of bacterial species estimated to exist on Earth has increased dramatically in recent years. This newly recognized species diversity has raised the possibility that bacterial natural product biosynthetic diversity has also been significantly underestimated by previous culture-based studies. Here, we compare 454-pyrosequenced nonribosomal peptide adenylation domain, type I polyketide ketosynthase domain, and type II polyketide ketosynthase alpha gene fragments amplified from cosmid libraries constructed using DNA isolated from three different arid soils. While 16S rRNA gene sequence analysis indicates these cloned metagenomes contain DNA from similar distributions of major bacterial phyla, we found that they contain almost completely distinct collections of secondary metabolite biosynthetic gene sequences. When grouped at 85% identity, only 1.5% of the adenylation domain, 1.2% of the ketosynthase, and 9.3% of the ketosynthase alpha sequence clusters contained sequences from all three metagenomes. Although there is unlikely to be a simple correlation between biosynthetic gene sequence diversity and the diversity of metabolites encoded by the gene clusters in which these genes reside, our analysis further suggests that sequences in one soil metagenome are so distantly related to sequences in another metagenome that they are, in many cases, likely to arise from functionally distinct gene clusters. The marked differences observed among collections of biosynthetic genes found in even ecologically similar environments suggest that prokaryotic natural product biosynthesis diversity is, like bacterial species diversity, potentially much larger than appreciated from culture-based studies.
Figures
References
-
- Ayuso-Sacido A, Genilloud O. 2005. New PCR primers for the screening of NRPS and PKS-I systems in actinomycetes: detection and distribution of these biosynthetic gene sequences in major taxonomic groups. Microb. Ecol. 49:10–24 - PubMed
-
- Beijerinck MW. 1913. De Infusies en de Ontdekking der Backterien. In van de Koninklijke Jaarboek, van Wetenschappen Akademie Müller, Amsterdam, The Netherlands
-
- Brady SF. 2007. Construction of soil environmental DNA cosmid libraries and screening for clones that produce biologically active small molecules. Nat. Protoc. 2:1297–1305 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
