

In vitro selection of highly modified cyclic peptides that act as tight binding inhibitors
- PMID: 22428867
- PMCID: PMC3384292
- DOI: 10.1021/ja301017y
In vitro selection of highly modified cyclic peptides that act as tight binding inhibitors
Abstract
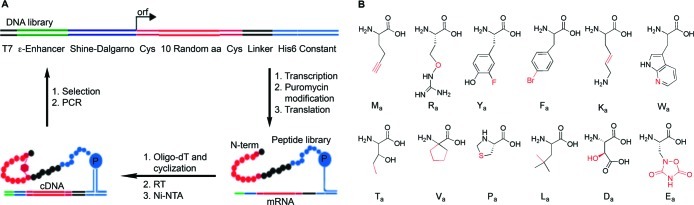
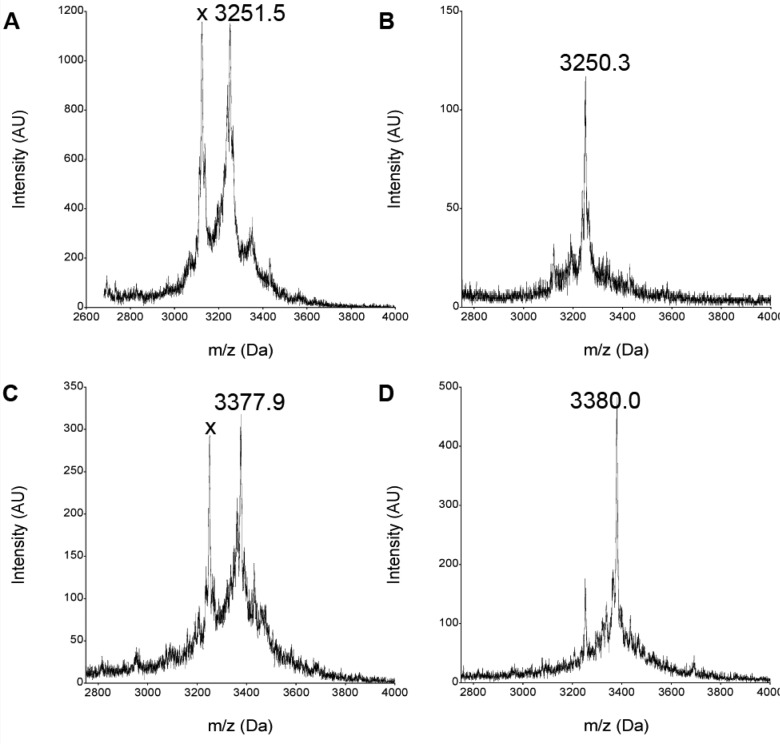
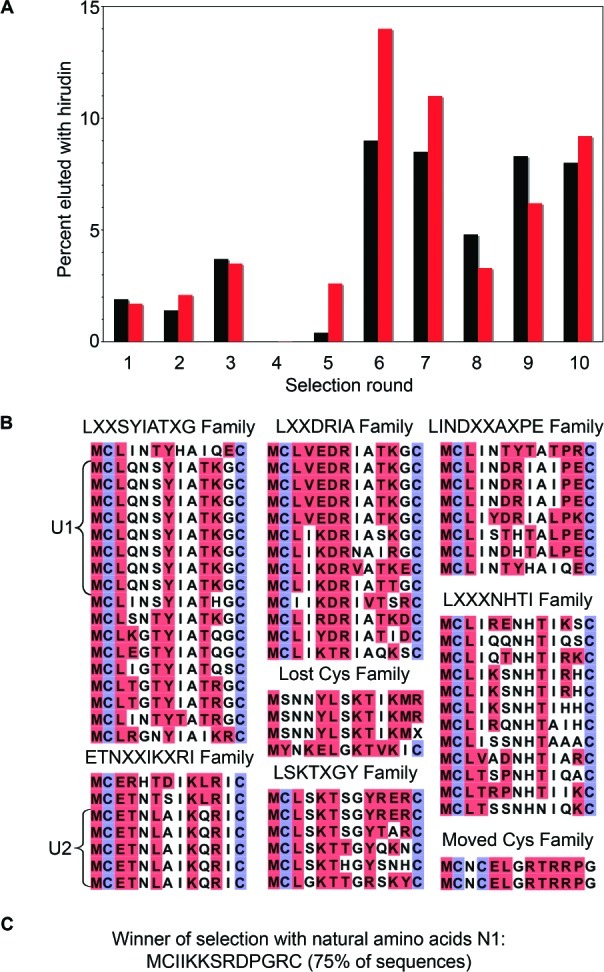
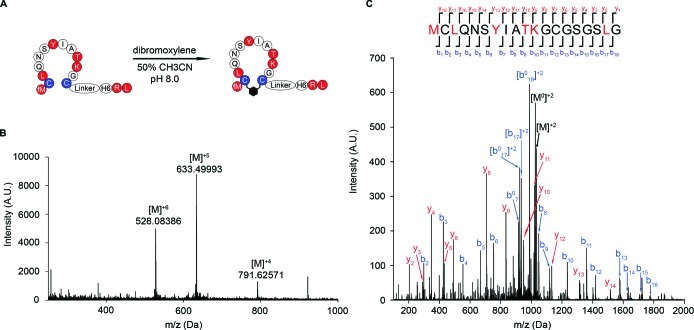
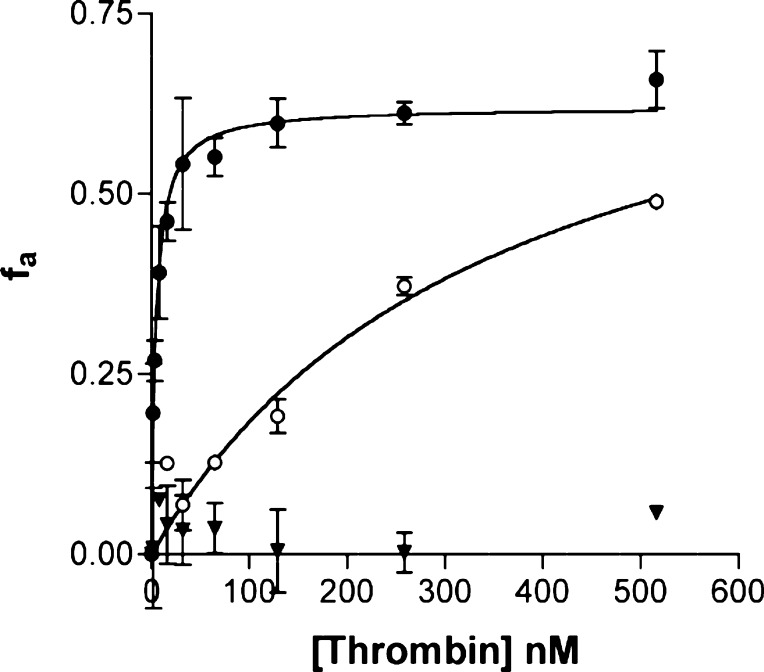
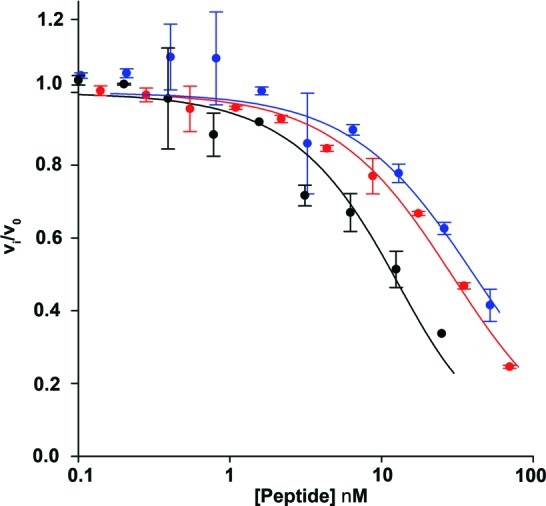
There is a great demand for the discovery of new therapeutic molecules that combine the high specificity and affinity of biologic drugs with the bioavailability and lower cost of small molecules. Small, natural-product-like peptides hold great promise in bridging this gap; however, access to libraries of these compounds has been a limitation. Since ribosomal peptides may be subjected to in vitro selection techniques, the generation of extremely large libraries (>10(13)) of highly modified macrocyclic peptides may provide a powerful alternative for the generation and selection of new useful bioactive molecules. Moreover, the incorporation of many non-proteinogenic amino acids into ribosomal peptides in conjunction with macrocyclization should enhance the drug-like features of these libraries. Here we show that mRNA-display, a technique that allows the in vitro selection of peptides, can be applied to the evolution of macrocyclic peptides that contain a majority of unnatural amino acids. We describe the isolation and characterization of two such unnatural cyclic peptides that bind the protease thrombin with low nanomolar affinity, and we show that the unnatural residues in these peptides are essential for the observed high-affinity binding. We demonstrate that the selected peptides are tight-binding inhibitors of thrombin, with K(i)(app) values in the low nanomolar range. The ability to evolve highly modified macrocyclic peptides in the laboratory is the first crucial step toward the facile generation of useful molecular reagents and therapeutic lead molecules that combine the advantageous features of biologics with those of small-molecule drugs.
Figures
References
-
- Verdine G. L.; Walensky L. D. Clin. Cancer Res. 2007, 13, 7264. - PubMed
-
- Reichert J. M.; Rosensweig C. J.; Faden L. B.; Dewitz M. C. Nat. Biotechnol. 2005, 23, 1073. - PubMed
-
- Malik N. N. Nat. Rev. Clin. Oncol. 2009, 6, 550. - PubMed
-
- Lipinski C. A. J. Pharmacol. Toxicol. Methods 2000, 44, 235. - PubMed
-
- Swinney D. C.; Anthony J. Nat. Rev. Drug Discov. 2011, 10, 507. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
